
 | ||
In molecular biology, splicing is the editing of the nascent precursor messenger RNA (pre-mRNA) transcript. After splicing, introns are removed and exons are joined together (ligated). For nuclear-encoded genes, splicing takes place within the nucleus either co-transcriptionally or immediately after transcription. For those eukaryotic genes that contain introns, splicing is usually required in order to create an mRNA molecule that can be translated into protein. For many eukaryotic introns, splicing is carried out in a series of reactions which are catalyzed by the spliceosome, a complex of small nuclear ribonucleoproteins (snRNPs). Self-splicing introns, or ribozymes capable of catalyzing their own excision from their parent RNA molecule, also exist.
Contents
Splicing pathways
Several methods of RNA splicing occur in nature; the type of splicing depends on the structure of the spliced intron and the catalysts required for splicing to occur.
Introns
The word intron is derived from the term intervening sequence, that is, a segment of DNA that "intervenes" between two exons of a gene. The term intron refers to both the DNA sequence within a gene and the corresponding sequence in the unprocessed RNA transcript. As part of the RNA processing pathway, introns are removed by RNA splicing either shortly after or concurrent with transcription. Introns are found in the genes of most organisms and many viruses. They can be located in a wide range of genes, including those that generate proteins, ribosomal RNA (rRNA), and transfer RNA (tRNA).
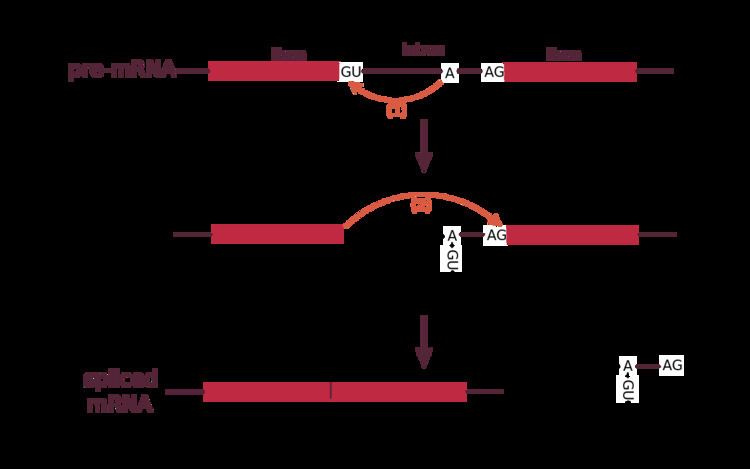
Spliceosomal introns often reside within the sequence of eukaryotic protein-coding genes. Within the intron, a donor site (5' end of the intron), a branch site (near the 3' end of the intron) and an acceptor site (3' end of the intron) are required for splicing. The splice donor site includes an almost invariant sequence GU at the 5' end of the intron, within a larger, less highly conserved region. The splice acceptor site at the 3' end of the intron terminates the intron with an almost invariant AG sequence. Upstream (5'-ward) from the AG there is a region high in pyrimidines (C and U), or polypyrimidine tract. Further upstream from the polypyrimidine tract is the branchpoint, which includes an adenine nucleotide involved in lariat formation. The consensus sequence for an intron (in IUPAC nucleic acid notation) is: G-G-[cut]-G-U-R-A-G-U (donor site) ... intron sequence ... Y-U-R-A-C (branch sequence 20-50 nucleotides upstream of acceptor site) ... Y-rich-N-C-A-G-[cut]-G (acceptor site). However, it is noted that the specific sequence of intronic splicing elements and the number of nucleotides between the branchpoint and the nearest 3’ acceptor site affect splice site selection. Also, point mutations in the underlying DNA or errors during transcription can activate a cryptic splice site in part of the transcript that usually is not spliced. This results in a mature messenger RNA with a missing section of an exon. In this way, a point mutation, which might otherwise affect only a single amino acid, can manifest as a deletion or truncation in the final protein.
Formation and activity
Splicing is catalyzed by the spliceosome, a large RNA-protein complex composed of five small nuclear ribonucleoproteins (snRNPs, pronounced 'snurps'). Assembly and activity of the spliceosome occurs during transcription of the pre-mRNA. The RNA components of snRNPs interact with the intron and are involved in catalysis. Two types of spliceosomes have been identified (major and minor) which contain different snRNPs.
Self-splicing
Self-splicing occurs for rare introns that form a ribozyme, performing the functions of the spliceosome by RNA alone. There are three kinds of self-splicing introns, Group I, Group II and Group III. Group I and II introns perform splicing similar to the spliceosome without requiring any protein. This similarity suggests that Group I and II introns may be evolutionarily related to the spliceosome. Self-splicing may also be very ancient, and may have existed in an RNA world present before protein.
Two transesterifications characterize the mechanism in which group I introns are spliced:
- 3'OH of a free guanine nucleoside (or one located in the intron) or a nucleotide cofactor (GMP, GDP, GTP) attacks phosphate at the 5' splice site.
- 3'OH of the 5' exon becomes a nucleophile and the second transesterification results in the joining of the two exons.
The mechanism in which group II introns are spliced (two transesterification reaction like group I introns) is as follows:
- The 3'OH of a specific adenosine in the intron attacks the 5' splice site, thereby forming the lariat
- The 3'OH of the 5' exon triggers the second transesterification at the 3' splice site, thereby joining the exons together.
tRNA splicing
tRNA (also tRNA-like) splicing is another rare form of splicing that usually occurs in tRNA. The splicing reaction involves a different biochemistry than the spliceosomal and self-splicing pathways.
In the yeast Saccharomyces cerevisiae, a yeast tRNA splicing endonuclease heterotetramer, composed of TSEN54, TSEN2, TSEN34, and TSEN15, cleaves pre-tRNA at two sites in the acceptor loop to form a 5'-half tRNA, terminating at a 2',3'-cyclic phosphodiester group, and a 3'-half tRNA, terminating at a 5'-hydroxyl group, along with a discarded intron. Yeast tRNA kinase then phosphorylates the 5'-hydroxyl group using adenosine triphosphate. Yeast tRNA cyclic phosphodiesterase cleaves the cyclic phosphodiester group to form a 2'-phosphorylated 3' end. Yeast tRNA ligase adds an adenosine monophosphate group to the 5' end of the 3'-half and joins the two halves together. NAD-dependent 2'-phosphotransferase then removes the 2'-phosphate group.
Evolution
Splicing occurs in all the kingdoms or domains of life, however, the extent and types of splicing can be very different between the major divisions. Eukaryotes splice many protein-coding messenger RNAs and some non-coding RNAs. Prokaryotes, on the other hand, splice rarely and mostly non-coding RNAs. Another important difference between these two groups of organisms is that prokaryotes completely lack the spliceosomal pathway.
Because spliceosomal introns are not conserved in all species, there is debate concerning when spliceosomal splicing evolved. Two models have been proposed: the intron late and intron early models (see intron evolution).
Biochemical mechanism
Spliceosomal splicing and self-splicing involve a two-step biochemical process. Both steps involve transesterification reactions that occur between RNA nucleotides. tRNA splicing, however, is an exception and does not occur by transesterification.
Spliceosomal and self-splicing transesterification reactions occur via two sequential transesterification reactions. First, the 2'OH of a specific branchpoint nucleotide within the intron, defined during spliceosome assembly, performs a nucleophilic attack on the first nucleotide of the intron at the 5' splice site, forming the lariat intermediate. Second, the 3'OH of the released 5' exon then performs a nucleophilic attack at the first nucleotide following the last nucleotide of the intron at the 3' splice site, thus joining the exons and releasing the intron lariat.
Alternative splicing
In many cases, the splicing process can create a range of unique proteins by varying the exon composition of the same mRNA. This phenomenon is then called alternative splicing. Alternative splicing can occur in many ways. Exons can be extended or skipped, or introns can be retained. It is estimated that 95% of transcripts from multiexon genes undergo alternative splicing, some instances of which occur in a tissue-specific manner and/or under specific cellular conditions. Development of high throughput mRNA sequencing technology can help quantify the expression levels of alternatively spliced isoforms. Differential expression levels across tissues and cell lineages allowed computational approaches to be developed to predict the functions of these isoforms. Given this complexity, alternative splicing of pre-mRNA transcripts is regulated by a system of trans-acting proteins (activators and repressors) that bind to cis-acting sites or "elements" (enhancers and silencers) on the pre-mRNA transcript itself. These proteins and their respective binding elements promote or reduce the usage of a particular splice site. The binding specificity comes from the sequence and structure of the cis-elements e.g. in HIV-1 there are many donor and acceptor splice sites. Among the various splice sites ssA7 which is 3' acceptor site folds into three stem loop structure i.e. Intronic splicing silencer (ISS), Exonic splicing enhancer (ESE) and Exonic splicing silencer (ESSE3). Solution structure of Intronic splicing silencer and its interaction to host protein hnRNPA1 give insight into specific recognition. However, adding to the complexity of alternative splicing, it is noted that the effects of regulatory factors are many times position-dependent. For example, a splicing factor that serves as a splicing activator when bound to an intronic enhancer element may serve as a repressor when bound to its splicing element in the context of an exon, and vice versa. In addition to the position-dependent effects of enhancer and silencer elements, the location of the branchpoint (i.e., distance upstream of the nearest 3’ acceptor site) also affects splicing. The secondary structure of the pre-mRNA transcript also plays a role in regulating splicing, such as by bringing together splicing elements or by masking a sequence that would otherwise serve as a binding element for a splicing factor.
Experimental manipulation of splicing
Splicing events can be experimentally altered by binding steric-blocking antisense oligos such as Morpholinos or Peptide nucleic acids to snRNP binding sites, to the branchpoint nucleotide that closes the lariat, or to splice-regulatory element binding sites.
Splicing errors and variation
It has been suggested that one third of all disease-causing mutations impact on splicing. Common errors include:
Although many splicing errors are safeguarded by a cellular quality control mechanism termed nonsense-mediated mRNA decay (NMD), a number of splicing-related diseases also exist, as suggested above.
Allelic differences in mRNA splicing are likely to be a common and important source of phenotypic diversity at the molecular level, in addition to their contribution to genetic disease susceptibility. Indeed, genome-wide studies in humans have identified a range of genes that are subject to allele-specific splicing.
Protein splicing
In addition to RNA, proteins can undergo splicing. Although the biomolecular mechanisms are different, the principle is the same: parts of the protein, called inteins instead of introns, are removed. The remaining parts, called exteins instead of exons, are fused together. Protein splicing has been observed in a wide range of organisms, including bacteria, archaea, plants, yeast and humans.