
Scientific name Pseudomonas syringae | ||
 | ||
Similar Pseudomonas, Bacteria, Xanthomonas, Erwinia, Thale cress | ||
The bacterial actinidia pseudomonas syringae pv actinidiae english version
Pseudomonas syringae is a rod-shaped, Gram-negative bacterium with polar flagella. As a plant pathogen, it can infect a wide range of species, and exists as over 50 different pathovars, all of which are available to researchers from international culture collections such as the NCPPB, ICMP, and others. Whether these pathovars represent a single species is unclear.
Contents
- The bacterial actinidia pseudomonas syringae pv actinidiae english version
- Bacterial rot of grapevine caused by pseudomonas syringae
- History
- Epidemiology
- Mechanisms of pathogenicity
- Ability to invade plants
- Overcoming host resistance
- Biofilm formation
- Ice nucleating properties
- Pathovars
- Determinants of host specificity
- P syringae as a model system
- P syringae pv tomato strain DC3000 and Arabidopsis thaliana
- References
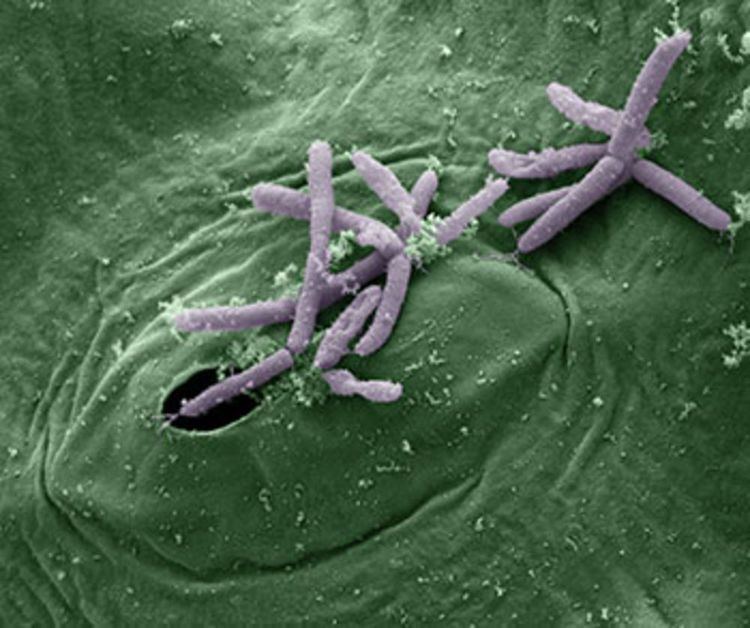
P. syringae is a member of the Pseudomonas genus, and based on 16S rRNA analysis, it has been placed in the P. syringae group. It is named after the lilac tree (Syringa vulgaris), from which it was first isolated.

P. syringae tests negative for arginine dihydrolase and oxidase activity, and forms the polymer levan on sucrose nutrient agar. Many, but not all, strains secrete the lipodepsinonapeptide plant toxin syringomycin, and it owes its yellow fluorescent appearance when cultured in vitro on King's B medium to production of the siderophore pyoverdin.

P. syringae also produces ice nucleation active (INA) proteins which cause water (in plants) to freeze at fairly high temperatures (-4 to -2°C), resulting in injury. Since the 1970s, P. syringae has been implicated as an atmospheric "biological ice nucleator", with airborne bacteria serving as cloud condensation nuclei. Recent evidence has suggested the species plays a larger role than previously thought in producing rain and snow. They have also been found in the cores of hailstones, aiding in Bioprecipitation. These INA proteins are also used in making artificial snow.
P. syringae pathogenesis is dependent on effector proteins secreted into the plant cell by the bacterial type III secretion system. Nearly 60 different type III effector families encoded by hop genes have been identified in P. syringae. Type III effectors contribute to pathogenesis chiefly through their role in suppressing plant defense. Owing to early availability of the genome sequence for three P. syringae strains and the ability of selected strains to cause disease on well-characterized host plants, including Arabidopsis thaliana, Nicotiana benthamiana, and the tomato, P. syringae has come to represent an important model system for experimental characterization of the molecular dynamics of plant-pathogen interactions.
Bacterial rot of grapevine caused by pseudomonas syringae
History
In 1961, Paul Hoppe of the U.S. Department of Agriculture studied a corn fungus by grinding up infected leaves each season, then applying the powder to test corn for the following season to track the disease. A surprise frost occurred that year, leaving peculiar results. Only plants infected with the diseased powder incurred frost damage, leaving healthy plants unfrozen. This phenomenon baffled scientists until graduate student Stephen Lindow of the University of Wisconsin–Madison with D.C. Arny and C. Upper found a bacterium in the dried leaf powder in the early 1970s. Dr. Lindow, now a plant pathologist at the University of California-Berkeley, found that when this particular bacterium was introduced to plants where it is originally absent, the plants became very vulnerable to frost damage. He went on to identify the bacterium as P. syringae, investigate P. syringae's role in ice nucleation and in 1977, discover the mutant ice-minus strain. He was later successful at developing the ice-minus strain of P. syringae through recombinant DNA technology, as well.
Epidemiology
Disease by P. syringae tends to be favoured by wet, cool conditions—optimum temperatures for disease tend to be around 12–25°C, although this can vary according to the pathovar involved. The bacteria tend to be seed-borne, and are dispersed between plants by rain splash.
Although it is a plant pathogen, it can also live as a saprotroph in the phyllosphere when conditions are not favourable for disease. Some saprotrophic strains of P. syringae have been used as biocontrol agents against postharvest rots.
Mechanisms of pathogenicity
The mechanisms of P. syringae pathogenicity can be separated into several categories: ability to invade a plant, ability to overcome host resistance, biofilm formation, and production of proteins with ice-nucleating properties.
Ability to invade plants
Planktonic P. syringae is able to enter plants using its flagella and pilli to swim towards a target host. It enters the plant via wounds of natural opening sites, as it is not able to breach the plant cell wall. The role of taxis in P. syringae has not been well-studied, but the bacteria are thought to use chemical signals released by the plant to find their host and cause infection.
Overcoming host resistance
P. syringae isolates carry a range of virulence factors called type III secretion system (T3SS) effector proteins. These proteins primarily function to cause disease symptoms and manipulate the host's immune response to facilitate infection. The major family of T3SS effectors in P. syringae is the hrp gene cluster, coding for the Hrp secretion apparatus.
The pathogens also produce phytotoxins which injure the plant and can suppress the host immune system. One such phytotoxin is coronatine, found in pathovars Pto and Pgl.
Biofilm formation
P. syringae produces polysaccharides which allow it to adhere to the surface of plant cells. It also releases quorum sensing molecules, which allows it to sense the presence of other bacterial cells nearby. If these molecules pass a threshold level, the bacteria change their pattern of gene expression to form a biofilm and begin expression virulence-related genes. The bacteria secrete highly viscous compounds such as polysaccharides and DNA to create a protective environment in which to grow.
Ice-nucleating properties
P. syringae—more than any mineral or other organism—is responsible for the surface frost damage in plants exposed to the environment. For plants without antifreeze proteins, frost damage usually occurs between -4 and -12°C as the water in plant tissue can remain in a supercooled liquid state. P. syringae can cause water to freeze at temperatures as high as −1.8°C (28.8°F), but strains causing ice nucleation at lower temperatures (down to −8 °C) are more common. The freezing causes injuries in the epithelia and makes the nutrients in the underlying plant tissues available to the bacteria.
P. syringae has ina (ice nucleation-active) genes that make INA proteins which translocate to the outer bacterial membrane on the surface of the bacteria, where the proteins act as nuclei for ice formation. Artificial strains of P. syringae known as ice-minus bacteria have been created to reduce frost damage.
P. syringae has been found in the center of hailstones, suggesting the bacterium may play a role in Earth's hydrological cycle.
Pathovars
Following ribotypical analysis, incorporation of several pathovars of P. syringae into other species was proposed (see P. amygdali, 'P. tomato', P. coronafaciens, P. avellanae, 'P. helianthi', P. tremae, P. cannabina, and P. viridiflava). According to this schema, the remaining pathovars are:
However, many of the strains for which new species groupings were proposed continue to be referred to in the scientific literature as pathovars of P. syringae, including pathovars tomato, phaseolicola, and maculicola. Pseudomonas savastanoi was once considered a pathovar or subspecies of P. syringae, and in many places continues to be referred to as P. s. pv. savastanoi, although as a result of DNA-relatedness studies, it has been instated as a new species. It has three host-specific pathovars: P. s. fraxini (which causes ash canker), P. s. nerii (which attacks oleander), and P. s. oleae (which causes olive knot).
Determinants of host specificity
A combination of the pathogen's effector genes and the plant's resistance genes is thought to determine which species a particular pathovar can infect. Plants can develop resistance to a pathovar by recognising pathogen-associated molecular patterns ((PAMPs)) and launching an immune response. These PAMPs are necessary for the microbe to function, so cannot be lost, but the pathogen may find ways to suppress this immune response, leading to an evolutionary arms race between the pathogen and the host.
P. syringae as a model system
Owing to early availability of genome sequences for P syringae pv, tomato strain DC3000, P. syringae pv. syringae strain B728a, and P. syringae pv. phaseolicola strain 1448A, together with the ability of selected strains to cause disease on well-characterized host plants such as Arabidopsis thaliana, Nicotiana benthamiana, and tomato, P. syringae has come to represent an important model system for experimental characterization of the molecular dynamics of plant-pathogen interactions. The P. syringae experimental system has been a source of pioneering evidence for the important role of pathogen gene products in suppressing plant defense. The nomenclature system developed for P. syringae effectors has been adopted by researchers characterizing effector repertoires in other bacteria, and methods used for bioinformatic effector identification have been adapted for other organisms. In addition, researchers working with P. syringae have played an integral role in the Plant-Associated Microbe Gene Ontology working group, aimed at developing gene ontology terms that capture biological processes occurring during the interactions between organisms, and using the terms for annotation of gene products.
P. syringae pv. tomato strain DC3000 and Arabidopsis thaliana
As mentioned above, the genome of P. syringae pv. tomato DC3000 has been sequenced, and approximately 40 Hop (Hrp Outer Protein) effectors, pathogenic proteins that attenuate the host cell, have been identified. These 40 effectors are not recognized by A. thaliana thus making P. syringae pv. tomato DC3000 virulent, that is, P. syringae pv. tomato DC3000 is able to infect A. thaliana which is susceptible to this pathogen.
Many gene-for-gene relationships have been identified using the two model organisms, P. syringae pv. tomato strain DC3000 and Arabidopsis. The gene-for-gene relationship describes the recognition of pathogenic avirulence (avr) genes by host resistance genes (R-genes). P. syringae pv. tomato DC3000 is a useful tool for studying avr: R-gene interactions in A. thaliana because it can be transformed with avr genes from other bacterial pathogens, and furthermore, because none of the endogenous hops genes is recognized by A. thaliana, any observed aver recognition identified using this model can be attributed to recognition of the introduced avr by A. thaliana. The transformation of P. syringae pv tomato DC3000 with effectors from other pathogens have led to the identification of many R-genes in Arabidopsis to further advance knowledge of plant pathogen interactions.