
Group Group I (dsDNA) | Rank Family | |
 | ||
Similar Bracovirus, Ascoviridae, Cotesia congregata, Cotesia, Phycodnaviridae | ||
Polydnavirus
A polydnavirus /pɒˈlɪdnəvaɪərəs/ (PDV) is a member of the family Polydnaviridae of insect viruses. There are currently 53 species in this family, divided among 2 genera. Polydnaviruses form a symbiotic relationship with parasitoid wasps (ichnoviruses occur in ichneumonid wasps species and bracoviruses in braconid wasps), but these wasps are themselves parasites of lepidoptera. Little or no sequence homology exists between BV and IV, suggesting that the two genera evolved independently.
Contents
- Polydnavirus
- How to pronounce polydnavirus
- Taxonomy
- Structure
- Life Cycle
- Biology
- Characteristics
- Evolution
- Effect on host immunity
- Virus like particles
- Micro RNA
- References
How to pronounce polydnavirus
Taxonomy
Group: dsDNA
Structure
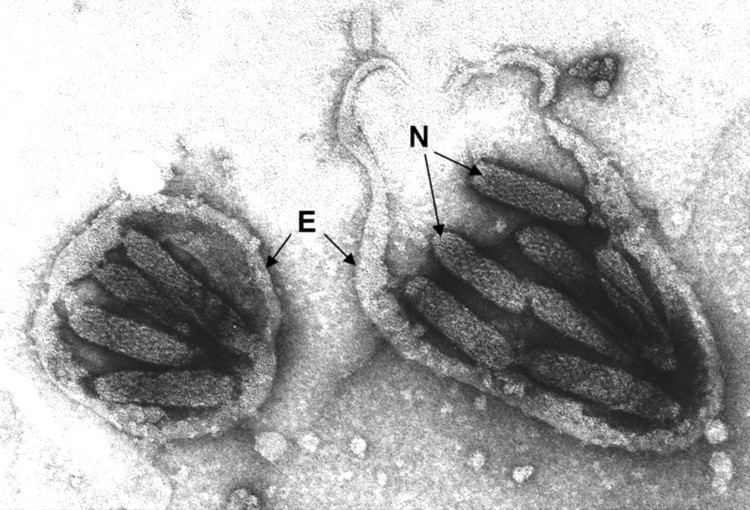
Viruses in Polydnaviridae are enveloped, with prolate ellipsoid and cylindrical geometries. Genomes are circular and segmented, composed of multiple segments of double-stranded, superhelical DNA packaged in capsid proteins. They are around 2.0–31kb in length.
Life Cycle
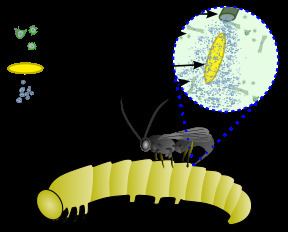
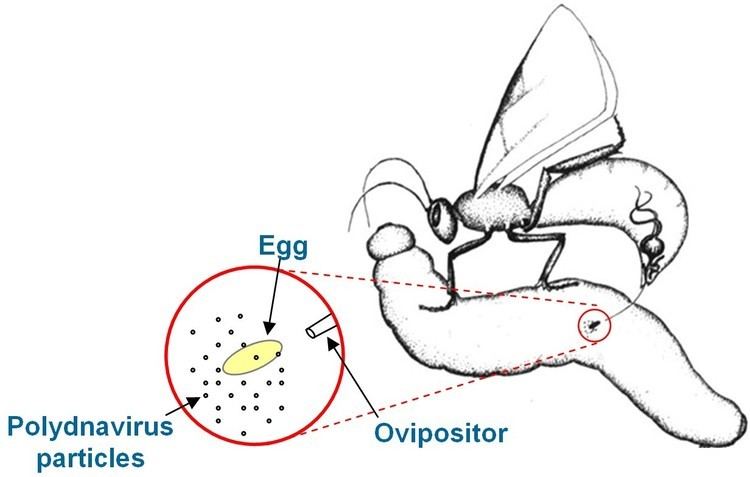
Viral replication is nuclear. DNA-templated transcription is the method of transcription. The virus exits the host cell by nuclear pore export. Parasitoid wasps serve as hosts, but these wasps are themselves parasites of lepidoptera. The wasp injects one or more eggs into its host along with a quantity of virus. The virus and wasp are in a symbiotic relationship: expression of viral genes prevents the wasp's host's immune system from killing the wasp's injected egg and causes other physiological alterations that ultimately cause the parasitized host to die. Transmission routes are parental.
Biology
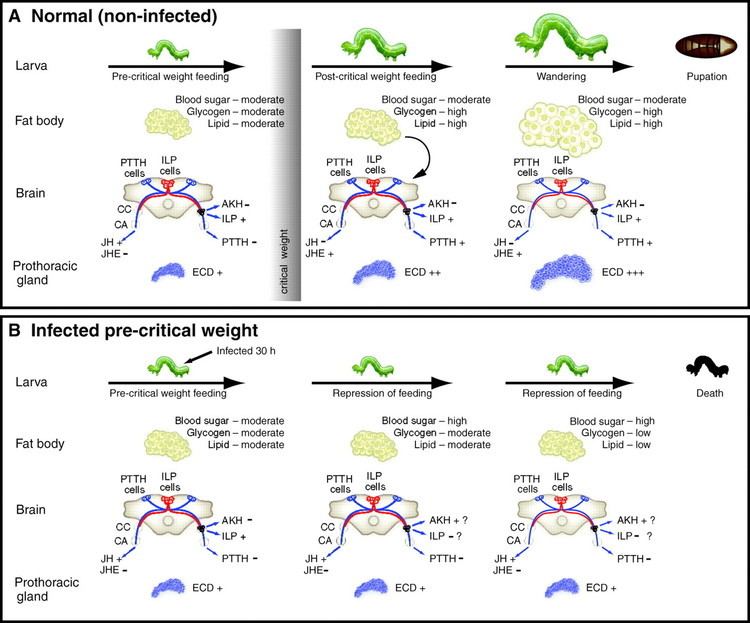
These viruses are part of a unique biological system consisting of an endoparasitic wasp (parasitoid), an insect (usually lepidopteran) larva, and the virus. The full genome of the virus is integrated into the genome of the wasp and the virus only replicates in a particular part of the ovary, called the calyx, of pupal and adult female wasps. The virus is injected along with the wasp egg into the body cavity of a lepidopteran host caterpillar and infects cells of the caterpillar. The infection does not lead to replication of new viruses, rather it affects the caterpillar's immune system. Without the virus infection, phagocytic hemocytes (blood cells) will encapsulate and kill the wasp egg but the immune suppression caused by the virus allows for survival of the wasp egg, leading to hatching and complete development of the immature wasp in the caterpillar. Additionally, genes expressed from the polydnavirus in the parasitised host alter host development and metabolism to be beneficial for the growth and survival of the parasitoid larva. Thus the virus and wasp have a symbiotic (mutualistic) relationship.
Characteristics
Both genera of PDV share certain characteristics:
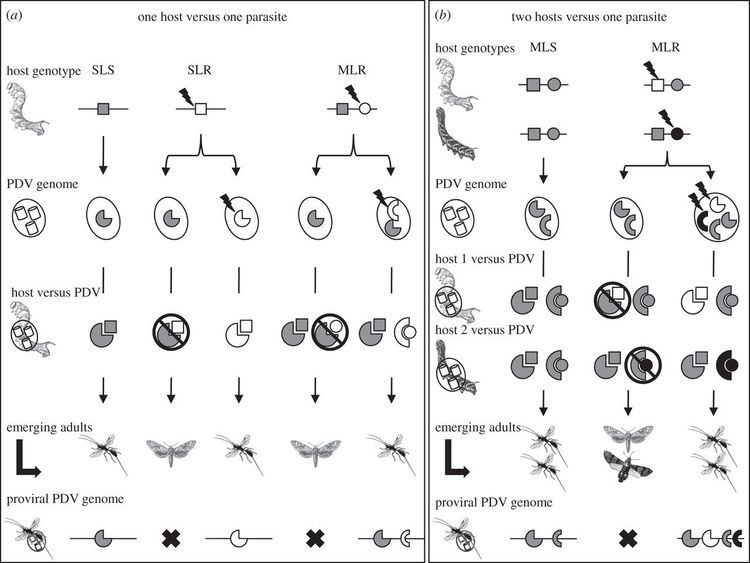
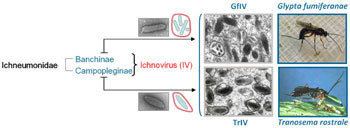
However, the morphology of the two genera are different when observed by electron microscopy. Ichnoviruses tend to be ovoid (egg-shaped) while bracoviruses are short rods. The virions of Bracoviruses are released by cell lysis whereas virions of Ichnoviruses are released by budding.
Evolution
Nucleic acid analysis suggests a very long association of the viruses with the wasps (greater than 70 million years).
Two proposals have been advanced for how the wasp/virus association developed. The first suggests that the virus is derived from wasp genes. Many parasitoids that do not use PDVs inject proteins that provide many of the same functions, that is, a suppression of the immune response to the parasite egg. In this model, the braconid and ichneumonid wasps packaged genes for these functions into the viruses—essentially creating a gene-transfer system that results in the caterpillar producing the immune-suppressing factors. In this scenario, the PDV structural proteins (capsids) were probably "borrowed" from existing viruses.
The alternative proposal suggests that ancestral wasps developed a beneficial association with an existing virus that eventually led to the integration of the virus into the wasp’s genome. Following integration, the genes responsible for virus replication and the capsids were (eventually) no longer included in the PDV genome. This hypothesis is supported by the distinct morphology differences between IV and BV, suggesting different ancestral viruses for the two families. The IV have remarkable similarities to ascoviruses while BV may have evolved from a nudivirus ~100 million years ago.
The two groups of viruses in the family are not in fact phylogenetically related suggesting that this taxon may need revision.
Effect on host immunity
In the host, several mechanisms of the insect immune system can be triggered when the wasp lays its eggs and when the parasitic wasp is developing.
When a large body (wasp egg or small particle used experimentally) is introduced into an insect's body, the classic immune reaction is the encapsulation by the hematocytes. An encapsulated body can also be melanised in order to asphyxiate it, thanks to another type of hemocyte, which uses the phenoloxidase pathway to produce melanin. Small particles can be phagocytosed, and macrophage cells can then be also melanised in a nodule. Finally, insects can also respond with production of antiviral peptides.
PolyDNAvirus protect the hymenopteran larvae from the host immune system, acting at different levels.
Virus like particles
Another strategy used by parasitoid Hymenoptera to protect their offspring is production of Virus Like Particles. VLPs are very close to viruses in their structure but they don't carry any nucleic acid. For example, Venturia canescens (Ichneumonidea) and Leptopilina sp. (Figitidaea) produce VLPs.
VLPs can be compared to PolyDNAvirus because they are secreted in the same way, and they both act to protect the larvae against the host's immune system. V. canescens-VLPs (VcVLP1, VcVLP2,VcNEP …) are produced in the calix cells before they go to the oviducts. But they are composed of proteins that are closer in structure and function to Hymenoptera's proteins than to viral proteins. This suggests that they don't have a viral origin, unlike polyDNAvirus.
VLPs protect the Hymenoptera larvae locally, whereas polyDNAvirus can have a more global effect. VLPs allow the larvae to escape the immune system: the larva is not recognised as harmful by its host, or the immune cells can't interact with it thanks to the VLPs.
The wasp Leptopilina heterotoma secrete VLPs that are able to penetrate into the lamellocytes, thanks to specific receptors, and then modify their shape and surface properties so they become inefficient and the larvae are safe from encapsulation.
Micro-RNA
MicroRNA are small RNA fragments produced in the host cells thanks to a specific enzymatic mechanism. They promote viral RNA destruction. MicroRNA attach to viral-RNA because they are complementary. Then the complex is recognised by an enzyme that destroys it. This phenomenon is known as PTGS (for post transcriptional gene silencing)
It is interesting to consider the microRNA phenomenon in the polyDNAvirus context. Many hypotheses can be formulated :