
Group Group II (ssDNA) Scientific name Parvoviridae | Order Unassigned Rank Family | |
 | ||
Lower classifications Parvovirus, Canine parvovirus, Feline distemper, Porcine parvovirus, Parvovirus B19 | ||
Parvoviridae mamma mia
The Parvoviridae are a family of small, rugged, genetically-compact DNA viruses, known collectively as parvoviruses. There are currently 56 species in the family, divided among 26 genera and two subfamilies. Members of this family infect a wide array of hosts and have been divided into two subfamilies, which infect either vertebrates (the Parvovirinae) or invertebrates (Densovirinae).
Contents
- Parvoviridae mamma mia
- Micro vet 2015 2 aula 4 fam lia parvoviridae
- Structure
- Genome
- Summary
- Attachment and Entry
- Single stranded DNA to double stranded DNA
- Transcription and translation
- DNA replication
- Taxonomy
- Viruses infecting humans
- Unclassified viruses
- Additional genera
- References
Parvovirus B19 was the first pathogenic human parvovirus to be discovered and is best known for causing a childhood exanthem called "fifth disease" (erythema infectiosum), although it is also associated with other diseases including arthritis.
Micro vet 2015 2 aula 4 fam lia parvoviridae
Structure
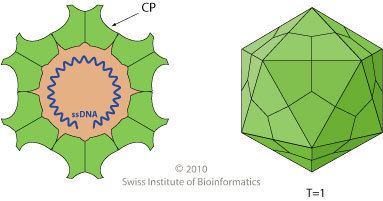
Parvovirus particles (virions) have a durable non-enveloped protein capsid ~20–30 nm in diameter that contains a single copy of the linear single-stranded ~ 5kb DNA genome, which terminates in small imperfect palindromes that fold into dynamic hairpin telomeres. These terminal hairpins are hallmarks of the family, giving rise to the viral origins of DNA replication and mediating multiple steps in the viral life cycle including genome amplification, packaging, and the establishment of transcription complexes. However, they are often refractory to detection by PCR amplification strategies since they tend to induce polymerase strand-switching. Many parvoviruses are exceptionally resistant to inactivation, remaining infectious for months or years after release into the environment.
Viruses in this family have small protein virions that exhibit T=1 icosahedral symmetry. Their capsid shells are assembled from 60 icosahedrally-ordered copies of a single core protein (VP) sequence, but some of these VP proteins also have N-terminal extensions that are not visible in X-ray structures. Biochemical and serological studies indicate that these extensions become successively exposed at the particle surface during virus maturation and cell entry, where they contribute to virion stability and mediate specific steps in cell trafficking. Parvoviruses appear to be unique in encoding a broad spectrum phospholipase A2 (PLA2) activity, typically in the N-terminus of the longest (VP1) subset of their capsid proteins, which is deployed to mediate virion transfer across the lipid bilayer of host cells
Genome
The viral genome is 4–6 kilobases in length and terminates in imperfectly-palindromic hairpin sequences of ~120–500 nucleotides that exhibit genus-specific secondary structures, and can either be identical at the two ends of the genome (homotelomeric) or can differ in size, sequence and predicted secondary structure (heterotelomeric). Homotelomeric viruses package DNA strands of both senses (into separate capsids) whereas heterotelomeric viruses generally package predominantly negative-sense DNA (discussed in references 5 and 6). All parvoviruses encode two major gene complexes: the non-structural (or rep) gene that encodes the replication initiator protein (called NS1 or Rep), and the VP (or cap) gene, which encodes a nested set of ~2–6 size variants derived from the C-terminus of the single VP protein sequence. Members of the Parvovirinae also encode a few (1–4) small genus-specific ancillary proteins that are variably distributed throughout the genome, show little sequence homology to each other, and appear to serve an array of different functions in each genus (references 2–6). Viruses in most genera are mono-sense, meaning that both viral genes are transcribed in a single direction from open reading frames in the same (positive-sense) DNA strand, but members of one genus of homotelomeric invertebrate viruses (genus Ambidensovirus) show ambisense organization, with the NS and capsid proteins being transcribed in opposite directions from the 5’-ends of the two complementary DNA strands (see reference 1 and ).
The major non-structural protein, NS1, is a site- and strand-specific endonuclease belonging to the HuH protein superfamily, and also carries a AAA+ SF3 helicase domain. NS1 initiates and drives the viral “rolling hairpin” replication mechanism (RHR), which is a linear adaptation of the more-common “rolling-circle” replication strategy used by many small circular prokaryotic and viral replicons. RHR is a unidirectional mechanism that displaces a single, continuous DNA strand, which rapidly folds and refolds to generate a series of concatemeric duplex replication intermediates. Unit length genomes are then excised from these intermediates by the NS1 endonuclease (reviewed in references 5 and 6), and packaged 3’-to-5’ into preformed empty capsids driven by the SF3 helicase activity of NS1/Rep.
Summary
Viral replication is nuclear. Entry into the host cell is achieved by attachment to host receptors, which mediate internalization via endocytosis. Capsids are metastable, undergoing a series of structural shifts during cell entry that sequentially expose peptides carrying PLA2 activity and trafficking signals . These signals ultimately mediate delivery of the intact virion into the cell nucleus, where genome uncoating allows the establishment of viral DNA replication and transcription complexes that rely predominantly upon the synthetic machinery of their host cell. Replication follows the unidirectional strand displacement mechanism discussed above. Packaged virions from viruses in at least two genera (Protoparvovirus and Bocaparvovirus) have mechanisms that allow mature virions to be trafficked out of viable host cells prior to cell lysis, but members of most other genera are only released into the environment following death and lysis of the infected cell. Natural animal hosts for parvoviruses include a wide range of vertebrates, insects and some crustacea. For viruses in the Parvovirinae transmission routes are commonly fecal-oral or respiratory.
Attachment and Entry
A virion attaches to receptors on the surface of a potential host cell. In the case of B19 virus the host cell is a red blood cell precursor and the receptor is the blood group P antigen. The virion enters the cell by endocytosis and is released from the endosome into the cytoplasm, where it associates with microtubules and is transported to a nuclear pore. With a diameter of 18–26 nm, the parvovirus virion is small enough to pass through a nuclear pore, unlike the herpesvirus nucleocapsid, though there is evidence that the virion must undergo some structural changes before it can be transported into the nucleus. Nuclear localization signals have been found in the capsid proteins of some parvoviruses.
Single-stranded DNA to double-stranded DNA
In the nucleus the single-stranded virus genome is converted to dsDNA by a cell DNA polymerase. The ends of the genome are double stranded as a result of base pairing, and at the 3end the –OH group acts as a primer to which the enzyme binds.
Transcription and translation
The cell RNA polymerase II transcribes the virus genes and cell transcription factors play key roles. The primary transcript(s) undergo various splicing events to produce two size classes of mRNA. The larger mRNAs encode the non-structural proteins and the smaller mRNAs encode the structural proteins. The non-structural proteins are phosphorylated and play roles in the control of gene expression and in DNA replication.
DNA replication
After and virion assembly conversion of the ssDNA genome to dsDNA, the DNA is replicated by a mechanism called rolling-hairpin replication. This is a leading strand mechanism and sets parvoviruses apart from other DNA viruses, which replicate their genomes through leading and lagging strand synthesis. Procapsids are constructed from the structural proteins and each is filled by a copy of the virus genome, either a ( + ) DNA or a ( − ) DNA as appropriate. One of the non-structural proteins functions as a helicase to unwind the dsDNA so that a single strand can enter the procapsid.
Taxonomy
Group: ssDNA
This taxonomy was last updated by the International Committee on Virus Taxonomy (ICTV) Parvoviridae Study Group early in 2014, as detailed in references 1 and 12. The family is divided into two subfamilies: Parvovirinae, which infect vertebrates and Densovirinae, which infect invertebrates. Each subfamily has been subdivided into several genera.
Subfamily Densovirinae:
Subfamily Parvovirinae:
To date, very few viruses from the Densovirinae have been studied and sequenced, so the above taxonomy may poorly reflect the true diversity of this subfamily. Currently, all recognized members of genus Ambidensovirus, Brevidensovirus and Iterodensovirus infect insects, while the hepan- and penstyldensoviruses infect decapod shrimp. However, recently two viruses have been isolated that segregate phylogenetically with members of genus Ambidensovirus but infect non-insect hosts. One of these, called cherax quadricarinatus densovirus (CqDV), has been shown to infect and kill the freshwater crayfish Cherax quadricarinatus, subphylum Crustacea, while the other, called sea-star-associated densovirus (SSaDV), has been shown to replicate in and kill sea-stars and perhaps some other members of phylum Echinodermata.
Viruses infecting humans
Currently, viruses that infect humans are recognized in 5 genera: Bocaparvovirus (human bocavirus 1–4, HboV1–4), Dependoparvovirus (adeno-associated virus 1–5, AAV1–5), Erythroparvovirus (parvovirus B19, B19), Protoparvovirus (Bufavirus 1–2, BuV1–2) and Tetraparvovirus (human parvovirus 4 G1–3, PARV4 G1–3).
Unclassified viruses
Since databases contain vast numbers of sequences that might be considered parvoviral in origin but are not real viruses, the Parvoviridae Study group cite the following criteria (references 1 and 12) that must be established before a new viral sequence can be considered for recognition in the family: "In order for an agent to be classified in the family Parvoviridae, it must be judged to be an authentic parvovirus on the basis of having been isolated and sequenced or, failing this, on the basis of having been sequenced in tissues, secretions, or excretions of unambiguous host origin, supported by evidence of its distribution in multiple individual hosts in a pattern that is compatible with dissemination by infection. The sequence must be in one piece, contain all the nonstructural (NS) and virus particle (VP) coding regions, and meet the size constraints and motif patterns typical of the family."
This means that partial coding sequences or sequences from a single host animal, with no evidence of virus exposure in the rest of the population, will not be considered sufficiently validated. Samples from feces are particularly problematic because they may be derived from food and so do not have an unequivocal host animal. Similarly, samples from aquatic animals that are also present in the surrounding environment are difficult to attribute until they can be shown to directly infect other members of the presumed host species. Despite these caveats, every year many new viruses are identified that will likely merit recognition, so that the published taxonomy always trails studies in the field.
For example, a candidate parvovirus has recently been isolated by sequencing a histocytic sarcoma in a slow loris (Nycticebus coucang). The relationship between the virus and the sarcoma was not clear.
Additional genera
Proposed additional genera include Chapparvovirus and Marinoparvovirus. Confirmation of the genera awaits a decision by the ICTV.