
Group Group II (ssDNA) Genus Inovirus Higher classification Inovirus | Family Inoviridae Scientific name Enterobacteria phage M13 Rank Species | |
 | ||
Similar Enterobacteria phage T4, T7 phage, Lambda phage, Phi X 174, Escherichia | ||
Release of m13 bacteriophage
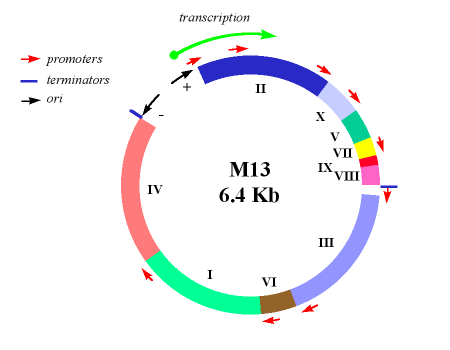
M13 is a virus that infects the bacterium Escherichia coli. It is composed of a circular single-stranded DNA molecule encased in a thin flexible tube made up of about 2700 copies of a single protein called P8, the major coat protein. The ends of the tube are capped with minor coat proteins. Infection starts when the minor coat protein P3 attaches to the receptor at the tip of the F pilus of the bacterium. Infection with M13 is not lethal; however, the infection causes turbid plaques in E. coli because infected bacteria grow more slowly than the surrounding uninfected bacteria. It engages in a viral lifestyle known as a chronic infection which is neither lytic or temperate. However a decrease in the rate of cell growth is seen in the infected cells. M13 plasmids are used for many recombinant DNA processes, and the virus has also been studied for its uses in nanostructures and nanotechnology.
Contents
Phage particles
The phage coat is primarily assembled from a 50 amino acid protein called pVIII (or p8), which is encoded by gene VIII (or g8) in the phage genome. For a wild type M13 particle, it takes approximately 2700 copies of p8 to make the coat about 900 nm long. The coat's dimensions are flexible though and the number of p8 copies adjusts to accommodate the size of the single stranded genome it packages. For example, when the phage genome was mutated to reduce its number of DNA bases (from 6.4 knt to 221 nt), then the number of p8 copies was decreased to fewer than 100, causing the p8 coat to shrink in order to fit the reduced genome. The phage appear to be limited at approximately twice the natural DNA content. However, deletion of a phage protein (p3) prevents full escape from the host E. coli, and phage that are 10-20X the normal length with several copies of the phage genome can be seen shedding from the E. coli host.
There are four other proteins on the phage surface, two of which have been extensively studied. At one end of the filament are five copies of the surface exposed pIX (p9) and a more buried companion protein, pVII (p7). If p8 forms the shaft of the phage, p9 and p7 form the "blunt" end that is seen in the micrographs. These proteins are very small, containing only 33 and 32 amino acids respectively, though some additional residues can be added to the N-terminal portion of each which are then presented on the outside of the coat. At the other end of the phage particle are five copies of the surface exposed pIII (p3) and its less exposed accessory protein, pVI (p6). These form the rounded tip of the phage and are the first proteins to interact with the E. coli host during infection. p3 is also the last point of contact with the host as new phage bud from the bacterial surface.
Replication in E. coli
Below are steps involved with replication of M13 in E. coli.
Phage proteins in the cytoplasm are pII, pX, and pV, and they are part of the replication process of DNA. The other phage proteins are synthesized and inserted into the cytoplasmic or outer membranes.
Applications
George Smith, among others, showed that fragments of EcoRI endonuclease could be fused in the unique Bam site of f1 filamentous phage and thereby expressed in gene III whose protein pIII was externally accessible. M13 does not have this unique Bam site in gene III. M13 had to be engineered to have accessible insertion sites, making it limited in its flexibility in handling different sized inserts. Because the M13 phage display system allows great flexibility in the location and number of recombinant proteins on the phage, it is a popular tool to construct or serve as a scaffold for nanostructures. For example, the phage can be engineered to have a different protein on each end and along its length. This can be used to assemble structures like gold or cobalt oxide nano-wires for batteries or to pack carbon nanotubes into straight bundles for use in photovoltaics.