
Group Group I (dsDNA) Higher classification Lambda-like viruses | Order Caudovirales Genus λ-like viruses Rank Species | |
 | ||
Scientific name Enterobacteria phage lambda Similar Enterobacteria phage T4, M13 bacteriophage, T7 phage, Escherichia, P1 phage | ||
Lambda phage part 1 the basic lifecycle
Enterobacteria phage λ (lambda phage, coliphage λ) is a bacterial Virus, or bacteriophage, that infects the bacterial species Escherichia coli (E. coli). It was discovered by Esther Lederberg in 1950 when she noticed that streaks of mixtures of two E. coli strains, one of which treated with ultraviolet light, was "nibbled and plaqued". This virus has a temperate lifecycle that allows it to either reside within the genome of its host through lysogeny or enter into a lytic phase (during which it kills and lyses the cell to produce offspring).
Contents
- Lambda phage part 1 the basic lifecycle
- Anatomy
- Infection
- N antitermination
- Lytic life cycle
- Rightward transcription
- Leftward transcription
- Lysogenic or lysenogenic life cycle
- Prophage integration
- Maintenance of lysogeny
- Induction
- Multiplicity reactivation and prophage reactivation
- Repressor
- Lytic or lysogenic
- Lambda as a genetic tool
- References
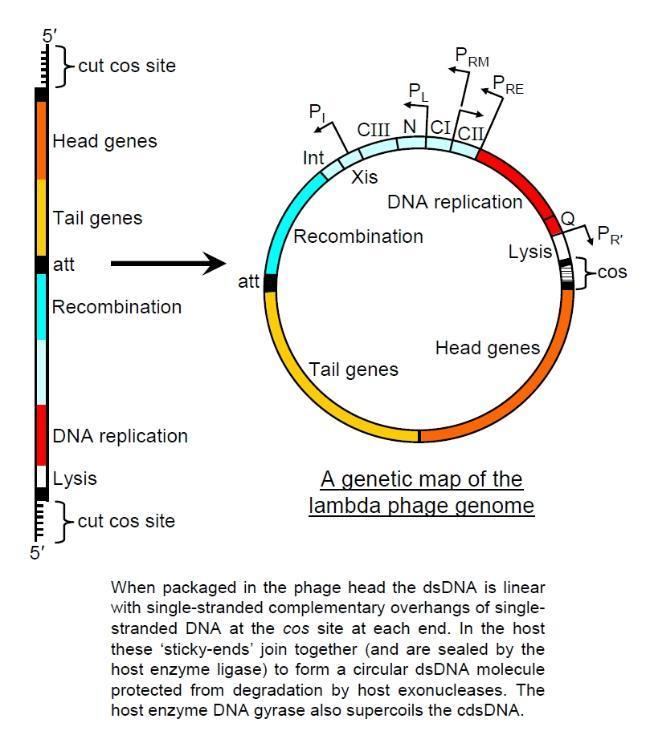
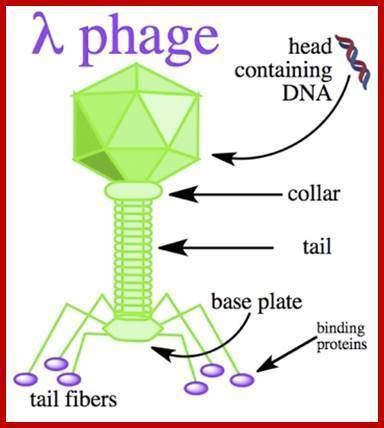
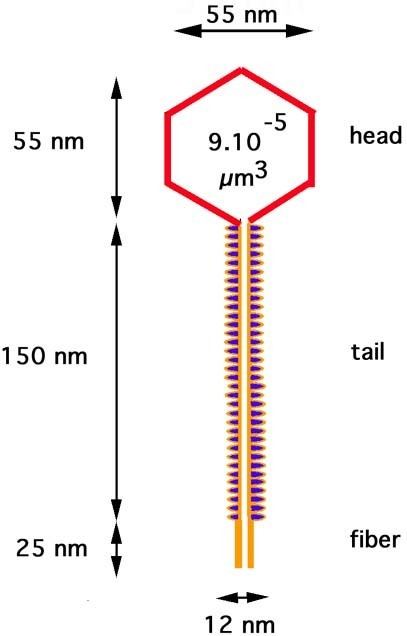
The phage particle consists of a head (also known as a capsid), a tail, and tail fibers (see image of virus below). The head contains the phage's double-strand linear DNA genome. During infection, the phage particle recognizes and binds to its host, E. coli, causing DNA in the head of the phage to be ejected through the tail into the cytoplasm of the bacterial cell. Usually, a "lytic cycle" ensues, where the lambda DNA is replicated and new phage particles are produced within the cell. This is followed by cell lysis, releasing the cell contents, including virions that have been assembled, into the environment. However, under certain conditions, the phage DNA may integrate itself into the host cell chromosome in the lysogenic pathway. In this state, the λ DNA is called a prophage and stays resident within the host's genome without apparent harm to the host. The host is termed a lysogen when a prophage is present. This prophage may enter the lytic cycle when the lysogen enters a stressed condition.
Anatomy
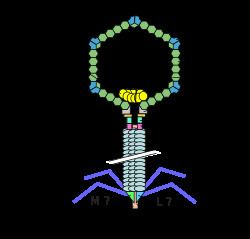
The virus particle consists of a head and a tail that can have tail fibers. The whole particle consists of 12–14 different proteins with more than 1000 protein molecules total and one DNA molecule located in the phage head. However, it is still not entirely clear whether the L and M proteins are part of the virion.
The genome contains 48,490 base pairs of double-stranded, linear DNA, with 12-base single-strand segments at both 5' ends. These two single-stranded segments are the "sticky ends" of what is called the cos site. The cos site circularizes the DNA in the host cytoplasm. In its circular form, the phage genome, therefore, is 48,502 base pairs in length. The lambda genome can be inserted into the E. coli chromosome and is then called a prophage. See section below for details.
Infection
Lambda phage is a non-contractile tailed phage, meaning during an infection event it cannot 'force' its DNA through a bacterial cell membrane. It must instead use an existing pathway to invade the host cell, having evolved the tip of its tail to interact with a specific pore to allow entry of its DNA to the hosts.
- Bacteriophage Lambda binds to an E. coli cell by means of its J protein in the tail tip. The J protein interacts with the maltose outer membrane porin (the product of the lamB gene) of E. coli, a porin molecule, which is part of the maltose operon.
- The linear phage genome is injected through the outer membrane.
- The DNA passes through the mannose permease complex in the inner membrane (encoded by the manXYZ genes) and immediately circularises using the cos sites, 12-base G-C-rich cohesive "sticky ends". The single-strand viral DNA ends are ligated by host DNA ligase.
- Host DNA gyrase puts negative supercoils in the circular chromosome, causing A-T-rich regions to unwind and drive transcription.
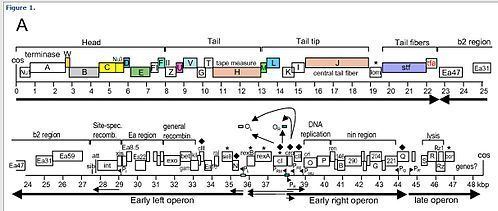
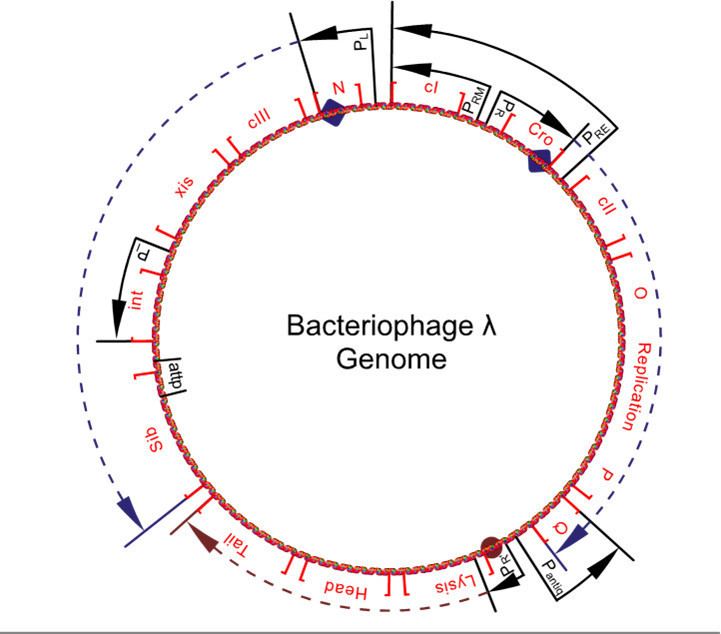
- Transcription starts from the constitutive PL, PR and PR' promoters producing the 'immediate early' transcripts. At first, these express the N and cro genes, producing N, Cro and a short inactive protein.
- Cro binds to OR3, preventing access to the PRM promoter, preventing expression of the cI gene. N binds to the two Nut (N utilisation) sites, one in the N gene in the PL reading frame, and one in the cro gene in the PR reading frame.
- The N protein is an antiterminator, and functions to extend the reading frames to which it is bound. When RNA polymerase transcribes these regions, it recruits the N and forms a complex with several host Nus proteins. This complex skips through most termination sequences. The extended transcripts (the 'late early' transcripts) include the N and cro genes along with cII and cIII genes, and xis, int, O, P and Q genes discussed later.
- The cIII protein acts to protect the cII protein from proteolysis by FtsH (a membrane-bound essential E. coli protease) by acting as a competitive inhibitor. This inhibition can induce a bacteriostatic state, which favours lysogeny. cIII also directly stabilises the cII protein.
On initial infection, the stability of cII determines the lifestyle of the phage; stable cII will lead to the lysogenic pathway, whereas if cII is degraded the phage will go into the lytic pathway. Low temperature, starvation of the cells and high multiplicity of infection (MOI) are known to favor lysogeny (see later discussion).
N antitermination
This occurs without the N protein interacting with the DNA; the protein instead binds to the freshly transcribed mRNA. Nut sites contain 3 conserved "boxes," of which only BoxB is essential.
- The boxB RNA sequences are located close to the 5' end of the pL and pR transcripts. When transcribed, each sequence forms a hairpin loop structure that the N protein can bind to.
- N protein binds to boxB in each transcript, and contacts the transcribing RNA polymerase via RNA looping. The N-RNAP complex is stabilized by subsequent binding of several host Nus (N utilisation substance) proteins (which include transcription termination/antitermination factors and, bizarrely, a ribosome subunit).
- The entire complex (including the bound Nut site on the mRNA) continues transcription, and can skip through termination sequences.
Lytic life cycle
This is the lifecycle that the phage follows following most infections, where the cII protein does not reach a high enough concentration due to degradation, so does not activate its promoters.
- The 'late early' transcripts continue being written, including xis, int, Q and genes for replication of the lambda genome (OP). Cro dominates the repressor site (see "Repressor" section), repressing synthesis from the PRM promoter (which is a promoter of the lysogenic cycle).
- The O and P proteins initiate replication of the phage chromosome (see "Lytic Replication").
- Q, another antiterminator, binds to Qut sites.
- Transcription from the PR' promoter can now extend to produce mRNA for the lysis and the head and tail proteins.
- Structural proteins and phage genomes self-assemble into new phage particles.
- Products of the lysis genes S,R, Rz and Rz1 cause cell lysis. S is a holin, a small membrane protein that, at a time determined by the sequence of the protein, suddenly makes holes in the membrane. R is an endolysin, an enzyme that escapes through the S holes and cleaves the cell wall. Rz and Rz1 are membrane proteins that form a complex that somehow destroys the outer membrane, after the endolysin has degraded the cell wall. For wild-type lambda, lysis occurs at about 50 minutes after the start of infection and releases around 100 virions.
Rightward transcription
Rightward transcription expresses the O, P and Q genes. O and P are responsible for initiating replication, and Q is another antiterminator that allows the expression of head, tail, and lysis genes from PR’.
Lytic replication
- For the first few replication cycles, the lambda genome undergoes θ replication (circle-to-circle).
- This is initiated at the ori site located in the O gene. O protein binds the ori site, and P protein binds the DnaB subunit of the host replication machinery as well as binding O. This effectively commandeers the host DNA polymerase.
- Soon, the phage switches to a rolling circle replication similar to that used by phage M13. The DNA is nicked and the 3’ end serves as a primer. Note that this does not release single copies of the phage genome but rather one long molecule with many copies of the genome: a concatemer.
- These concatemers are cleaved at their cos sites as they are packaged. Packaging cannot occur from circular phage DNA, only from concatomeric DNA.
Q antitermination
Q is similar to N in its effect: Q binds to RNA polymerase in Qut sites and the resulting complex can ignore terminators, however the mechanism is very different; the Q protein first associates with a DNA sequence rather than an mRNA sequence.
- The Qut site is very close to the PR’ promoter, close enough that the σ factor has not been released from the RNA polymerase holoenzyme. Part of the Qut site resembles the -10 Pribnow box, causing the holoenzyme to pause.
- Q protein then binds and displaces part of the σ factor and transcription re-initiates.
- The head and tail genes are transcribed and the corresponding proteins self-assemble.
Leftward transcription
Leftward transcription expresses the gam, red, xis, and int genes. Gam and red proteins are involved in recombination. Gam is also important in that it inhibits the host RecBCD nuclease from degrading the 3’ ends in rolling circle replication. Int and xis are integration and excision proteins vital to lysogeny.
xis and int regulation of insertion and excision
- xis and int are found on the same piece of mRNA, so approximately equal concentrations of xis and int proteins are produced. This results (initially) in the excision of any inserted genomes from the host genome.
- The mRNA from the PL promoter forms a stable secondary structure with a stem-loop in the sib section of the mRNA. This targets the 3' (sib) end of the mRNA for RNAaseIII degradation, which results in a lower effective concentration of int mRNA than xis mRNA (as the int cistron is nearer to the sib sequence than the xis cistron is to the sib sequence), so a higher concentrations of xis than int is observed.
- Higher concentrations of xis than int result in no insertion or excision of phage genomes, the evolutionarily favoured action - leaving any pre-insterted phage genomes inserted (so reducing competition) and preventing the insertion of the phage genome into the genome of a doomed host.
Lysogenic (or lysenogenic) life cycle
The lysogenic lifecycle begins once the cII protein reaches a high enough concentration to activate its promoters, after a small number of infections.
- The 'late early' transcripts continue being written, including xis, int, Q and genes for replication of the lambda genome.
- The stabilized cII acts to promote transcription from the PRE, PI and Pantiq promoters.
- The Pantiq promoter produces antisense mRNA to the Q gene message of the PR promoter transcript, thereby switching off Q production. The PRE promoter produces antisense mRNA to the cro section of the PR promoter transcript, turning down cro production, and has a transcript of the cI gene. This is expressed, turning on cI repressor production. The PI promoter expresses the int gene, resulting in high concentrations of int protein. This int protein integrates the phage DNA into the host chromosome (see "Prophage Integration").
- No Q results in no extension of the PR' promoter's reading frame, so no lytic or structural proteins are made. Elevated levels of int (much higher than that of xis) result in the insertion of the lambda genome into the hosts genome (see diagram). Production of cI leads to the binding of cI to the OR1 and OR2 sites in the PR promoter, turning off cro and other early gene expression. cI also binds to the PL promoter, turning off transcription there too.
- Lack of cro leaves the OR3 site unbound, so transcription from the PRM promoter may occur, maintaining levels of cI.
- Lack of transcription from the PL and PR promoters leads to no further production of cII and cIII.
- As cII and cIII concentrations decrease, transcription from the Pantiq, PRE and PI stop being promoted since they are no longer needed.
- Only the PRM and PR' promoters are left active, the former producing cI protein and the latter a short inactive transcript. The genome remains inserted into the host genome in a dormant state.
The prophage is duplicated with every subsequent cell division of the host. The phage genes expressed in this dormant state code for proteins that repress expression of other phage genes (such as the structural and lysis genes) in order to prevent entry into the lytic cycle. These repressive proteins are broken down when the host cell is under stress, resulting in the expression of the repressed phage genes. Stress can be from starvation, poisons (like antibiotics), or other factors that can damage or destroy the host. In response to stress, the activated prophage is excised from the DNA of the host cell by one of the newly expressed gene products and enters its lytic pathway.
Prophage integration
The integration of phage λ takes place at a special attachment site in the bacterial and phage genomes, called attλ. The sequence of the bacterial att site is called attB, between the gal and bio operons, and consists of the parts B-O-B', whereas the complementary sequence in the circular phage genome is called attP and consists of the parts P-O-P'. The integration itself is a sequential exchange (see genetic recombination) via a Holliday junction and requires both the phage protein Int and the bacterial protein IHF (integration host factor). Both Int and IHF bind to attP and form an intasome, a DNA-protein-complex designed for site-specific recombination of the phage and host DNA. The original B-O-B' sequence is changed by the integration to B-O-P'-phage DNA-P-O-B'. The phage DNA is now part of the host's genome.
Maintenance of lysogeny
Induction
The classic induction of a lysogen involved irradiating the infected cells with UV light. Any situation where a lysogen undergoes DNA damage or the SOS response of the host is otherwise stimulated leads to induction.
- The host cell, containing a dormant phage genome, experiences DNA damage due to a high stress environment, and starts to undergo the SOS response.
- RecA (a cellular protein) detects DNA damage and becomes activated. It is now RecA*, a highly specific co-protease.
- Normally RecA* binds LexA (a transcription repressor), activating LexA auto-protease activity, which destroys LexA repressor, allowing production of DNA repair proteins. In lysogenic cells, this response is hijacked, and RecA* stimulates cI autocleavage. This is because cI mimics the structure of LexA at the autocleavage site.
- Cleaved cI can no longer dimerise, and loses its affinity for DNA binding.
- The PR and PL promoters are no longer repressed and switch on, and the cell returns to the lytic sequence of expression events (note that cII is not stable in cells undergoing the SOS response). There is however one notable difference.
Control of phage genome excision in induction
- The phage genome is still inserted in the host genome and needs excision for DNA replication to occur. The sib section beyond the normal PL promoter transcript is, however, no longer included in this reading frame (see diagram).
- No sib domain on the PL promoter mRNA results in no hairpin loop on the 3' end, and the transcript is no longer targeted for RNAaseIII degradation.
- The new intact transcript has one copy of both xis and int, so approximately equal concentrations of xis and int proteins are produced.
- Equal concentrations of xis and int result in the excision of the inserted genome from the host genome for replication and later phage production.
Multiplicity reactivation and prophage reactivation
Multiplicity reactivation (MR) is the process by which multiple viral genomes, each containing inactivating genome damage, interact within an infected cell to form a viable viral genome. MR was originally discovered with phage T4, but was subsequently found in phage λ (as well as in numerous other bacterial and mammalian viruses). MR of phage λ inactivated by UV light depends on the recombination function of either the host or of the infecting phage. Absence of both recombination systems leads to a loss of MR.
Survival of UV-irradiated phage λ is increased when the E. coli host is lysogenic for an homologous prophage, a phenomenon termed prophage reactivation. Prophage reactivation in phage λ appears to occur by a recombinational repair process similar to that of MR.
Repressor
The repressor found in the phage lambda is a notable example of the level of control possible over gene expression by a very simple system. It forms a 'binary switch' with two genes under mutually exclusive expression, as discovered by Barbara J. Meyer.
The lambda repressor gene system consists of (from left to right on the chromosome):
The lambda repressor is a self assembling dimer also known as the cI protein. It binds DNA in the helix-turn-helix binding motif. It regulates the transcription of the cI protein and the Cro protein.
The life cycle of lambda phages is controlled by cI and Cro proteins. The lambda phage will remain in the lysogenic state if cI proteins predominate, but will be transformed into the lytic cycle if cro proteins predominate.
The cI dimer may bind to any of three operators, OR1, OR2, and OR3, in the order OR1 = OR2 > OR3. Binding of a cI dimer to OR1 enhances binding of a second cI dimer to OR2, an effect called cooperativity. Thus, OR1 and OR2 are almost always simultaneously occupied by cI. However, this does not increase the affinity between cI and OR3, which will be occupied only when the cI concentration is high.
At high concentrations of cI, the dimers will also bind to operators OL1 and OL2 (which are over 2 kb downstream from the R operators). When cI dimers are bound to OL1, OL2, OR1, and OR2 a loop is induced in the DNA, allowing these dimers to bind together to form an octamer. This is a phenomenon called long-range cooperativity. Upon formation of the octamer, cI dimers may cooperatively bind to OL3 and OR3, repressing transcription of cI. This autonegative regulation ensures a stable minimum concentration of the repressor molecule and, should SOS signals arise, allows for more efficient prophage induction.
Lytic or lysogenic?
An important distinction here is that between the two decisions; lysogeny and lysis on infection, and continuing lysogeny or lysis from a prophage. The latter is determined solely by the activation of RecA in the SOS response of the cell, as detailed in the section on induction. The former will also be affected by this; a cell undergoing an SOS response will always be lysed, as no cI protein will be allowed to build up. However, the initial lytic/lysogenic decision on infection is also dependent on the cII and cIII proteins.
In cells with sufficient nutrients, protease activity is high, which breaks down cII. This leads to the lytic lifestyle. In cells with limited nutrients, protease activity is low, making cII stable. This leads to the lysogenic lifestyle. cIII appears to stabilize cII, both directly and by acting as a competitive inhibitor to the relevant proteases. This means that a cell "in trouble", i.e. lacking in nutrients and in a more dormant state, is more likely to lysogenise. This would be selected for because the phage can now lie dormant in the bacterium until it falls on better times, and so the phage can create more copies of itself with the additional resources available and with the more likely proximity of further infectable cells.
A full biophysical model for lambda's lysis-lysogeny decision remains to be developed. Computer modeling and simulation suggest that random processes during infection drive the selection of lysis or lysogeny within individual cells. However, recent experiments suggest that physical differences among cells, that exist prior to infection, predetermine whether a cell will lyse or become a lysogen.
Lambda as a genetic tool
Lambda phage has played a significant role as a model organism in the fields of microbial and molecular genetics. It has served as a valuable resource for developing tools utilized in these disciplines. Among its applications, Lambda phage has been employed as a vector for cloning recombinant DNA. Additionally, its site-specific recombinase, known as 'int,' has been used in the gateway method for shuffling cloned DNAs. Furthermore, the Red operon of Lambda phage, which includes the proteins Red alpha (also referred to as 'exo'), beta, and gamma, has been instrumental in the development of a DNA engineering technique known as recombineering. Lambda phage has also been pivotal in advancing the understanding of specialized transduction.