
Scientific name Hamamelidaceae Rank Family | ||
 | ||
Lower classifications | ||
Persian parrotia in the hamamelidaceae family growing vid
Hamamelidaceae, commonly referred to as the witch-hazel family, is a family of flowering plants in the order Saxifragales. The clade consists of shrubs and small trees positioned within the woody clade of the core Saxifragales. The earlier Cronquist system recognized Hamamelidaceae in the Hamamelidales order.
Contents
- Persian parrotia in the hamamelidaceae family growing vid
- Distribution
- Subfamilies
- Tribes
- Taxonomy
- Characteristics
- Anthers
- Pollen
- Flowers
- Breeding systems
- Genera
- References

Distribution

The Hamamelidaceae were widely distributed in the Northern Hemisphere during the Upper Cretaceous and early Tertiary. Quaternary glaciation across the Northern Hemisphere caused the extinction of numerous species and the restricted distribution of others. Hamamelidaceae were completely obliterated from Europe along with numerous other genera of plants that were unable to escape the ice sheets due to geography (the Mediterranean Sea and Alps forming barriers that did not exist in North America and Asia)

The largest subfamily, the Hamamelidoideae, is now distributed in North America and western and eastern Asia. The Hamamelidoideae subtribe Dicoryphinae is now restricted to the African (including Madagascar and Comores) and Australian continents. Disanthoideae and Rhodoleioideae are now restricted to southern China and the Caucasus region. Mytilarioideae is restricted to eastern Asia. Altingioideae is now restricted to eastern Asia and western Asia and North America between central Mexico and Belize.
Subfamilies

Hamamelidaceae contains 27-30 genera and 80-140 species distributed among five to six subfamilies. The subfamilies are Exbucklandioideae, Rhodoleioideae, Mytilarioideae, Disanthoideae, Hamamelidoideae, and Altingioideae, which has been elevated to a family Altingiaceae in some recent treatments. Many of the subfamilies are monotypic and the majority of the species lie within the Hamamelidoideae, which has 22 genera.
The number of genera within each subfamily is as follows:

Tribes
Subdivision of the five smaller subfamilies of Exbucklandioideae, Rhodoleioideae, Mytilarioideae, Disanthoideae and Altingioideae sensu Endress (1989) has not been considered necessary. However, the largest subfamily, Hamamelidoideae, has been subdivided once by Harms (1930) into five tribes and then revisited and restructured by Shulze-Menz (1964). Four tribes have been recognized since then sensu Endress (1989).
The Hamamelideae can be further divided into subtribes Dicoryphinae, which includes genera distributed only in the southern hemisphere, Loropetalinae, with genera restricted to the tropical regions of Asia, and Hamamelidinae, which has one genus (Hamamelis) distributed in Asia and North America.
Taxonomy
The phylogenetic relationships of the Hamamelidaceae subfamilies have been revisited several times since the first comprehensive classification of the family. Morphological and DNA studies have supported monophyly of the Hamamelidoideae and have recognized the separation of the Rhodoleioideae and Disanthoideae subfamily and newly erected Mytilarioideae.
The long-standing question of whether Altingioideae should be a separate family has been assessed and supported by morphological and molecular phylogenetic studies. However, the phylogenetic works of recent researchers still refer to Altingioideae as a subfamily.
The relationships between Exbucklandioideae and the other subfamilies have proven controversial. The unresolved monophyly of Exbucklandioideae and the clades of Disanthoideae, Rhodoleioideae, Exbucklandioideae or even Mytilarioideae being a sister clade to Hamamelidoideae may have been a result of differing DNA methodologies researchers have used to produce phylogenetic trees and the inclusion or exclusion of certain genera used as outgroups in their analyses. However, the sister relationship of Disanthoideae and Hamamelidoideae has been well supported, although some researchers do not support this. Strong support for making Altingioideae a family has been recognized by textbooks and the Angiosperm Phylogeny Group. Research continues to resolve the deep relationships of the subfamilies within the Hamamelidaceae by incorporating whole or fragmentary fossil evidence.
Characteristics
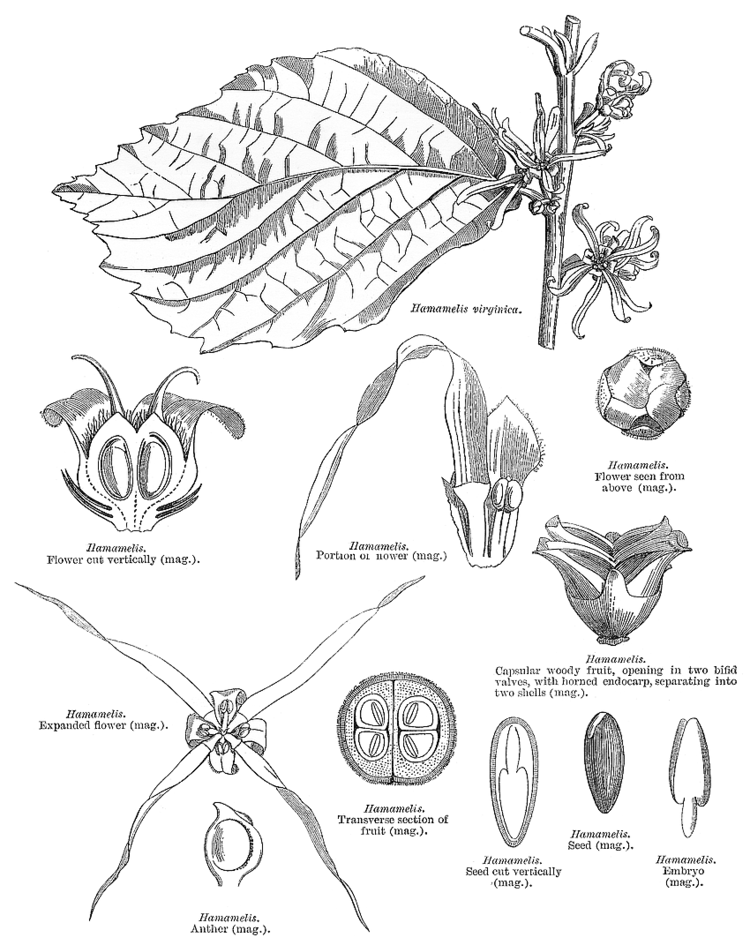
The Hamamelidaceae are distinguishable from other families in the Saxifragales due to the range of floral characteristics that are generally uniform though all genera. Uniform characteristics include stipules borne on stems with leaves often 2-ranked. Genera usually have a two carpel gynoecium, although some species show variation. Other characteristics include a multicellular stigma, with shallow papillae or ridges.
Anthers
Anther structure and the modes of opening are considered to be one of the most important features in the systematics and evolution of hamamelids. The anthers in Hamamelids are on average shorter than in other families in the Saxifragales. The anther valve openings are unique pleismorphic features that contrast with the simple longitudinal slits of the anthers in the upper Hamamelidae where the pollen is predominately wind-driven.
The three types of anthers found in the Hamamelidaceae are:
Pollen
Plants of the Hamamelidaceae have sticky pollen, which may have influenced the type of pollination that is seen in this family. Pollination is predominately via insects or wind. However, the insect-pollinated genus Disanthus has been known to wind-pollinate (although inefficiently) in the event pollinators do not visit its flowers. The genus Rhodoleia is unique because it is bird-pollinated
The pollen structure in the lower Hamamelidae is relatively uniform. The pollen patterns are tricolpate with reticulate exines.
Flowers
The petals of the Hamamelidaceae are generally narrow and ribbon-like. The exceptions are the genera Corylopsis and Rhodoleia, which have spathulate or circular-like petals. The flowers of Hamamelidaceae are mostly bisexual with perianth parts, which mature to fruits arranged in spikes, racemes or nonglobose heads.
Breeding systems
The anemophilous groups within the Hamamelidaceae are often andromonoecious. Self-incompatibility is common, but self-compatibility occurs in some genera such as Hamamelis.