
 | ||
In molecular biology and genetics, GC-content (or guanine-cytosine content) is the percentage of nitrogenous bases on a DNA or RNA molecule that are either guanine or cytosine (from a possibility of four different ones, also including adenine and thymine in DNA and adenine and uracil in RNA). This may refer to a certain fragment of DNA or RNA, or that of the whole genome. When it refers to a fragment of the genetic material, it may denote the GC-content of section of a gene (domain), single gene, group of genes (or gene clusters), or even a non-coding region. G (guanine) and C (cytosine) undergo a specific hydrogen bonding, whereas A (adenine) bonds specifically with T (thymine, in DNA) or U (uracil, in RNA).
Contents
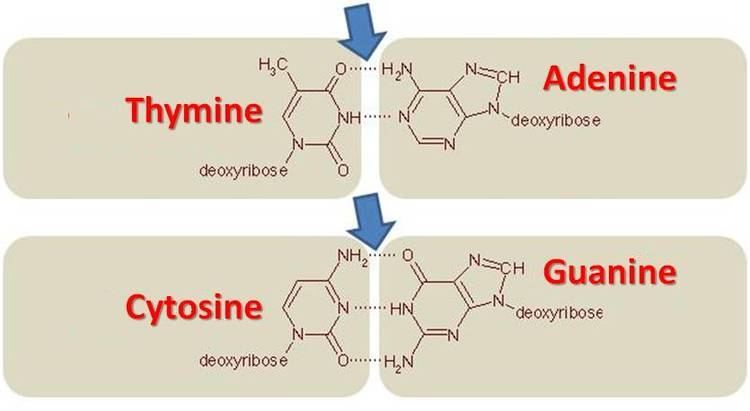
The GC pair is bound by three hydrogen bonds, while AT and AU pairs are bound by two hydrogen bonds. To emphasize this difference in the number of hydrogen bonds, the base pairings can be represented as respectively G≡C versus A=T and A=U. DNA with low GC-content is less stable than DNA high GC-content; however, the hydrogen bonds themselves do not have a particularly significant impact on stabilization, the stabilization is due mainly to interactions of base stacking. In spite of the higher thermostability conferred to the genetic material, it is envisaged that cells with DNA of high GC-content undergo autolysis, thereby reducing the longevity of the cell per se. Due to the thermostability given to the genetic materials in high GC organisms, it was commonly believed that the GC content played a necessary role in adaptation temperatures, a hypothesis that was refuted in 2001. However, it has been shown that there is a strong correlation between the prokaryotic optimal growth at higher temperatures and the GC content of structured RNAs (such as ribosomal RNA, transfer RNA, and many other non-coding RNAs). The AU base pairs are less stable than the GC base pairs because GC bonds have 3 hydrogen bonds and AU only has 2 hydrogen bonds, which makes high-GC-content RNA structures more resistant to the effects of high temperatures. More recently, one of the earliest large-scale systematic gene-centric association analysis recently demonstrated the correlation between GC content and temperature for certain genomic regions while not for others.
In PCR experiments, the GC-content of primers are used to predict their annealing temperature to the template DNA. A higher GC-content level indicates a higher melting temperature.
Determination
GC content is usually expressed as a percentage value, but sometimes as a ratio (called G+C ratio or GC-ratio). GC-content percentage is calculated as
whereas the AT/GC ratio is calculated as
The GC-content percentages as well as GC-ratio can be measured by several means, but one of the simplest methods is to measure what is called the melting temperature of the DNA double helix using spectrophotometry. The absorbance of DNA at a wavelength of 260 nm increases fairly sharply when the double-stranded DNA separates into two single strands when sufficiently heated. The most commonly used protocol for determining GC ratios uses flow cytometry for large number of samples.
In alternative manner, if the DNA or RNA molecule under investigation has been sequenced then the GC-content can be accurately calculated by simple arithmetic or by using the free online GC calculator.
GC ratio of genomes
GC ratios within a genome is found to be markedly variable. These variations in GC ratio within the genomes of more complex organisms result in a mosaic-like formation with islet regions called isochores. This results in the variations in staining intensity in the chromosomes. GC-rich isochores include in them many protein coding genes, and thus determination of ratio of these specific regions contributes in mapping gene-rich regions of the genome.
GC ratios and coding sequence
Within a long region of genomic sequence, genes are often characterised by having a higher GC-content in contrast to the background GC-content for the entire genome. Evidence of GC ratio with that of length of the coding region of a gene has shown that the length of the coding sequence is directly proportional to higher G+C content. This has been pointed to the fact that the stop codon has a bias towards A and T nucleotides, and, thus, the shorter the sequence the higher the AT bias.
Application in systematics
GC content is found to be variable with different organisms, the process of which is envisaged to be contributed to by variation in selection, mutational bias, and biased recombination-associated DNA repair. The species problem in prokaryotic taxonomy has led to various suggestions in classifying bacteria, and the ad hoc committee on reconciliation of approaches to bacterial systematics has recommended use of GC ratios in higher level hierarchical classification. For example, the Actinobacteria are characterised as "high GC-content bacteria". In Streptomyces coelicolor A3(2), GC content is 72%. The GC-content of Yeast (Saccharomyces cerevisiae) is 38%, and that of another common model organism, Thale Cress (Arabidopsis thaliana), is 36%. Because of the nature of the genetic code, it is virtually impossible for an organism to have a genome with a GC-content approaching either 0% or 100%. A species with an extremely low GC-content is Plasmodium falciparum (GC% = ~20%), and it is usually common to refer to such examples as being AT-rich instead of GC-poor.