
Scientific name Deuterostomia Rank Superphylum | ||
 | ||
Lower classifications Chordate, Vertebrate, Echinoderm, Reptile, Sea urchin | ||
Biology 2 lecture 12 deuterostomes
Deuterostomes (taxonomic term: Deuterostomia; meaning "second mouth" in Greek) are any members of a superphylum of animals. It is a sister clade of Protostomia, with which it forms the Nephrozoa clade.
Contents
- Biology 2 lecture 12 deuterostomes
- Protostomes and deuterostomes klt
- Classification history
- Notable characteristics
- Formation of mouth and anus
- Origins and evolution
- Classification
- References
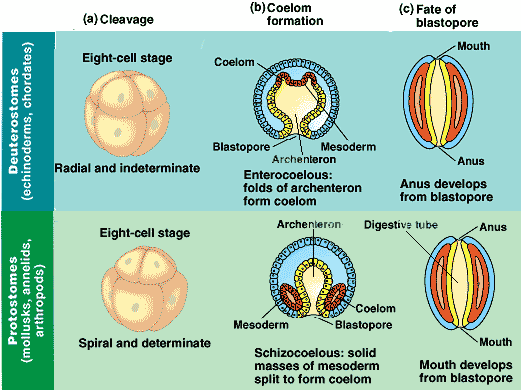
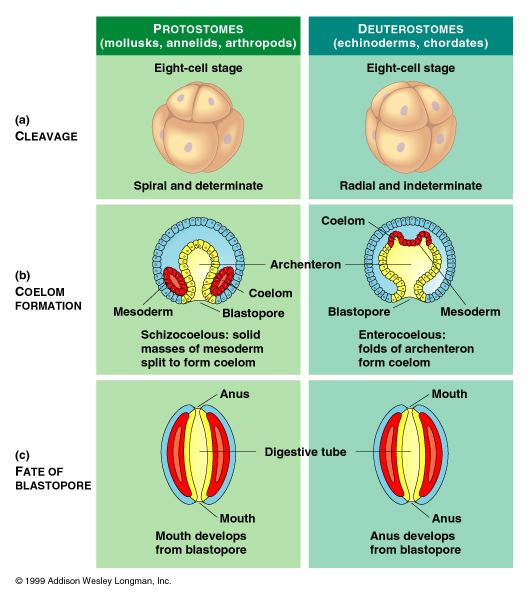
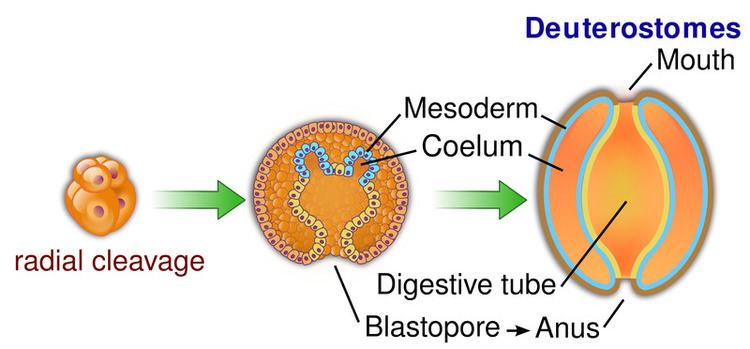
Deuterostomia is a subtaxon of the Bilateria branch of the subkingdom Eumetazoa, within Animalia, and are distinguished from protostomes by their deuterostomic embryonic development; in deuterostomes, the first opening (the blastopore) becomes the anus, while in protostomes, it becomes the mouth. (There are some occurrences of deuterostomy among Protostomes.)
Deuterostomes are also known as enterocoelomates because their coelom develops through enterocoely.
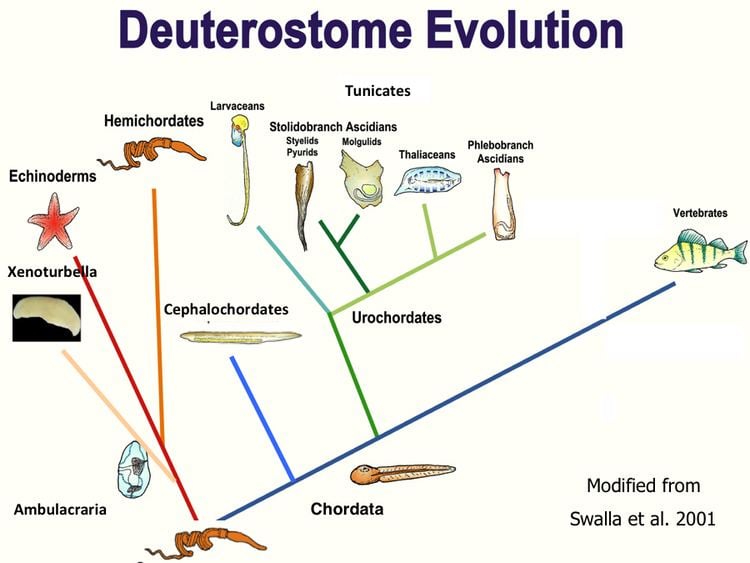
There are three major clades of deuterostomes:
Protostomes and deuterostomes klt
Classification history
Previously, Deuterostomia also included the phyla Brachiopoda, Bryozoa, Chaetognatha, and Phoronida based on embryological characteristics. However, Superphylum Deuterostomia was redefined in 1995 based on DNA molecular sequence analyses when the lophophorates were removed from it and combined with other protostome animals to form superphylum Lophotrochozoa. The phylum Chaetognatha (arrow worms) may belong here, but molecular studies have placed them in the protostomes more often.
Extinct deuterostome groups may include the phylum Vetulicolia.
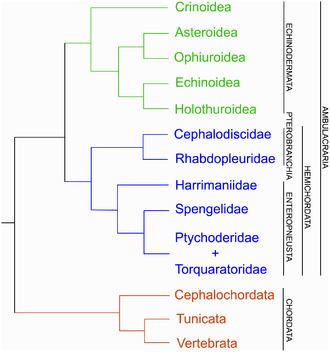
Echinodermata and Hemichordata form the clade Ambulacraria.
Notable characteristics
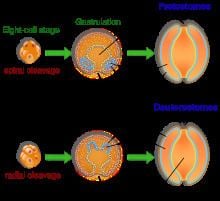
In both deuterostomes and protostomes, a zygote first develops into a hollow ball of cells, called a blastula. In deuterostomes, the early divisions occur parallel or perpendicular to the polar axis. This is called radial cleavage, and also occurs in certain protostomes, such as the lophophorates.
Most deuterostomes display indeterminate cleavage, in which the developmental fate of the cells in the developing embryo are not determined by the identity of the parent cell. Thus, if the first four cells are separated, each cell is capable of forming a complete small larva; and if a cell is removed from the blastula, the other cells will compensate.
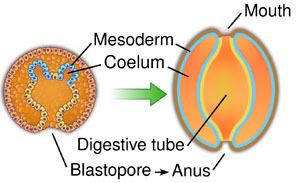
In deuterostomes the mesoderm forms as evaginations of the developed gut that pinch off, forming the coelom. This is called enterocoely.
Both the Hemichordata and Chordata have gill slits, and primitive fossil echinoderms also show signs of gill slits. A hollow nerve cord is found in all chordates, including tunicates (in the larval stage). Some hemichordates also have a tubular nerve cord. In the early embryonic stage, it looks like the hollow nerve cord of chordates.
Because of the highly modified nervous system of echinoderms, it is not possible to discern much about their ancestors in this matter, but based on different facts it is quite possible that all the present deuterostomes evolved from a common ancestor that had pharyngeal gill slits, a hollow nerve cord, circular and longitudinal muscles and a segmented body. It could have resembled the small group of Cambrian urochordate deuterostomes named Vetulicolia.
Formation of mouth and anus
The defining characteristic of the deuterostome is the fact that the blastopore (the opening at the bottom of the forming gastrula) becomes the anus, whereas in protostomes the blastopore becomes the mouth. The deuterostome mouth develops at the opposite end of the embryo from the blastopore and a digestive tract develops in the middle, connecting the two.
Origins and evolution
The majority of animals more complex than jellyfish and other Cnidarians are split into two groups, the protostomes and deuterostomes. Chordates (which include all the vertebrates) are deuterostomes.
It seems likely that the 555 million year old Kimberella was a member of the protostomes. That implies that the protostome and deuterostome lineages split some time before Kimberella appeared — at least 558 million years ago, and hence well before the start of the Cambrian 541 million years ago, i.e. during the later part of the Ediacaran Era (circa 635-542 Mya, around the end of global Marinoan glaciation in the late Neoproterozoic).
The oldest discovered proposed deuterostome is Saccorhytus coronarius, which lived approximately 540 million years ago. The researchers that made the discovery believe that the Saccorhytus is a common ancestor to all previously-known deuterostomes.
Fossils of one major deuterostome group, the echinoderms (whose modern members include sea stars, sea urchins and crinoids), are quite common from the start of Series 2 of the Cambrian, 521 million years ago. The Mid Cambrian fossil Rhabdotubus johanssoni has been interpreted as a pterobranch hemichordate. Opinions differ about whether the Chengjiang fauna fossil Yunnanozoon, from the earlier Cambrian, was a hemichordate or chordate. Another Chengjiang fossil, Haikouella lanceolata, also from the Chengjiang fauna, is interpreted as a chordate and possibly a craniate, as it shows signs of a heart, arteries, gill filaments, a tail, a neural chord with a brain at the front end, and possibly eyes — although it also had short tentacles round its mouth. Haikouichthys and Myllokunmingia, also from the Chengjiang fauna, are regarded as fish. Pikaia, discovered much earlier but from the Mid Cambrian Burgess Shale, is also regarded as a primitive chordate.
On the other hand, fossils of early chordates are very rare, as non-vertebrate chordates have no bone tissue or teeth, and fossils of no Post-Cambrian non-vertebrate chordates are known aside from the Permian-aged Paleobranchiostoma, trace fossils of the Ordovician colonial tunicate Catellocaula, and various Jurassic-aged and Tertiary-aged spicules tentatively attributed to ascidians.
Classification
Here are the following phyla/subgroups of the deuterostomes.