
Kingdom Animalia Scientific name Bilateria | Higher classification Eumetazoa | |
 | ||
Clade BilateriaHatschek, 1888 Lower classifications Vertebrate, Deuterostome, Protostome, Chordate, Arthropod | ||
The Bilateria /ˌbaɪləˈtɪəriə/ or bilaterians, or triploblasts, are animals with bilateral symmetry, i.e., they have a head ("anterior") and a tail ("posterior") as well as a back ("dorsal") and a belly ("ventral"); therefore they also have a left side and a right side. In contrast, radially symmetrical animals like jellyfish have a topside and a downside, but no identifiable front or back.
Contents
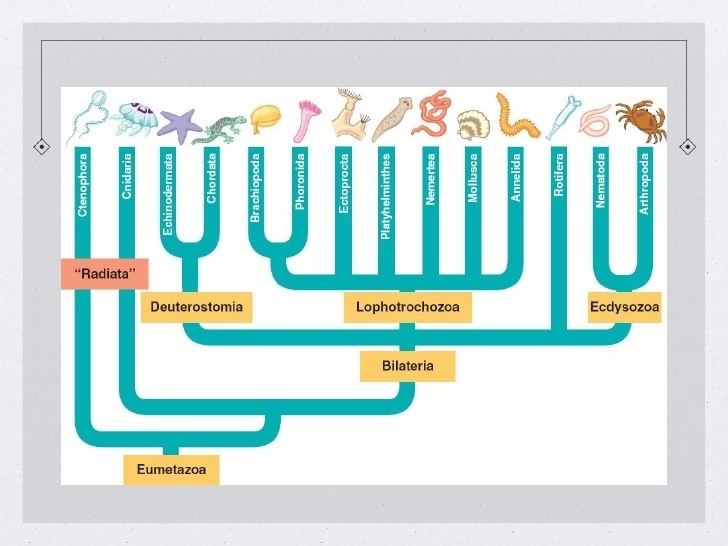
The bilateria are a major group of animals, including the majority of phyla but not sponges, cnidarians, placozoans and ctenophores. For the most part, bilateral embryos are triploblastic, having three germ layers: endoderm, mesoderm, and ectoderm. Nearly all are bilaterally symmetrical, or approximately so; the most notable exception is the echinoderms, which achieve near-radial symmetry as adults, but are bilaterally symmetrical as larvae.
Except for a few phyla (i.e. flatworms and gnathostomulids), bilaterians have complete digestive tracts with a separate mouth and anus. Some bilaterians lack body cavities (acoelomates, i.e. Platyhelminthes, Gastrotricha and Gnathostomulida), while others display primary body cavities (deriving from the blastocoel, as pseudocoel) or secondary cavities (that appear de novo, for example the coelom).
Bilateria
Evolution
The hypothetical most recent common ancestor of all bilateria is termed the "Urbilaterian". The nature of the first bilaterian is a matter of debate. One side suggests that acoelomates gave rise to the other groups (planuloid-aceloid hypothesis by Graff, Metchnikoff, Hyman, or Salvini Plawen), while the other poses that the first bilaterian was a coelomate organism and the main acoelomate phyla (flatworms and gastrotrichs) have lost body cavities secondarily (the Archicoelomata hypothesis and its variations such as the Gastrea by Haeckel or Sedgwick, the Bilaterosgastrea by Gösta Jägersten (sv.), or the Trochaea by Nielsen).
The first evidence of bilateria in the fossil record comes from trace fossils in Ediacaran sediments, and the first bona fide bilaterian fossil is Kimberella, dating to 555 million years ago. Earlier fossils are controversial; the fossil Vernanimalcula may be the earliest known bilaterian, but may also represent an infilled bubble. Fossil embryos are known from around the time of Vernanimalcula (580 million years ago), but none of these have bilaterian affinities. Burrows believed to have been created by bilaterian life forms have been found in the Tacuarí Formation of Uruguay, and are believed to be at least 585 million years old.
Phylogeny
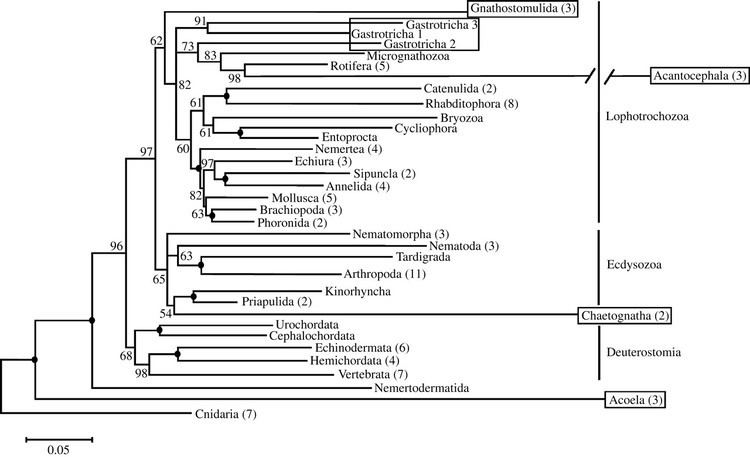
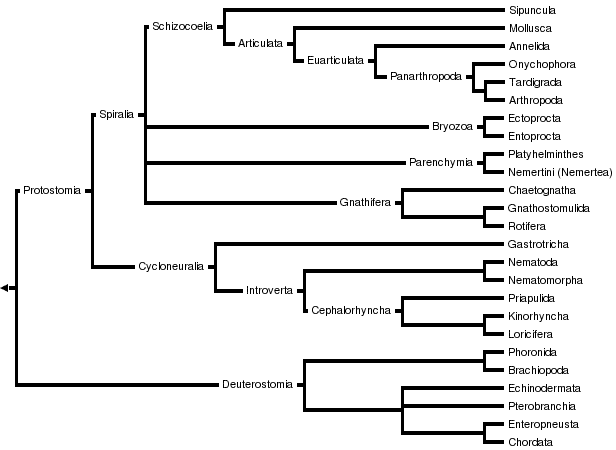
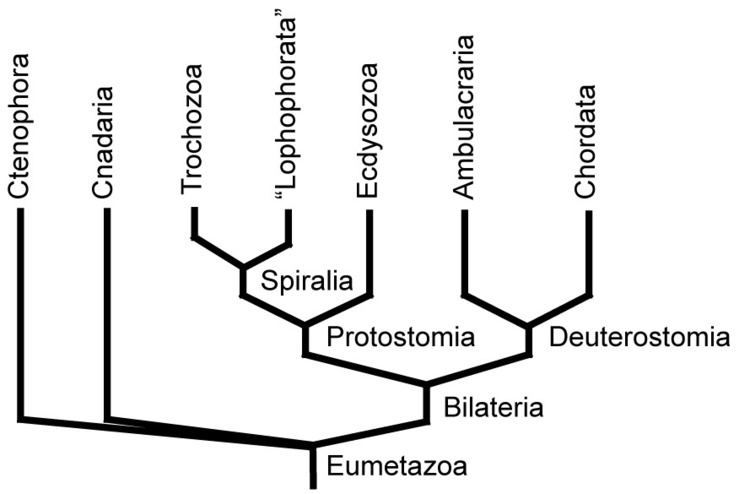
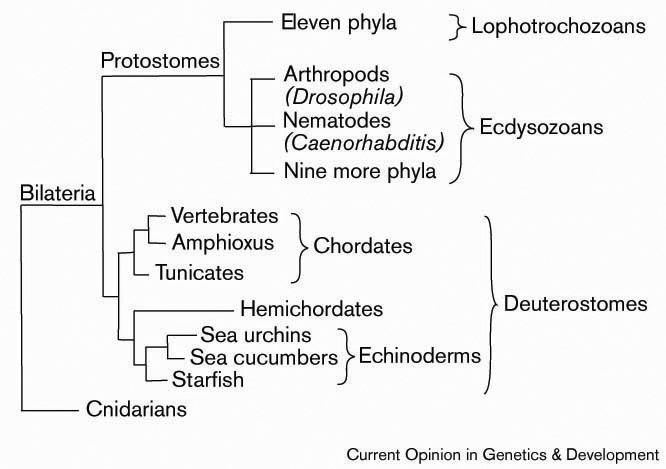
There are two main superphyla (main lineages) of Bilateria. The deuterostomes include the echinoderms, hemichordates, chordates, and a few smaller phyla. The protostomes include most of the rest, such as arthropods, annelids, mollusks, flatworms, and so forth. There are a number of differences, most notably in how the embryo develops. In particular, the first opening of the embryo becomes the mouth in protostomes, and the anus in deuterostomes. Many taxonomists now recognize at least two more superphyla among the protostomes, Ecdysozoa (molting animals) and Lophotrochozoa (also referred to as Spiralia). Within the latter, some researchers also recognize another superphylum, Platyzoa, while others reject the Platyzoa monophyly. The arrow worms (Chaetognatha) have proven particularly difficult to classify, with some taxonomists placing them among the deuterostomes and others placing them among the protostomes. The two most recent studies to address the question of chaetognath origins support protostome affinities.
A modern (2011) consensus phylogeny for Bilateria is shown below, although the position of certain clades are still controversial and the tree has changed considerably between 2000 and 2010. Nodes marked with * have received broad consensus. A prominent alternative tree is championed by Nielsen (2001).