
EC number 4.6.1.1 ExPASy NiceZyme view | CAS number 9012-42-4 | |
 | ||
Camp and adenylyl cyclase
Adenylyl cyclase (EC 4.6.1.1, also commonly known as adenyl cyclase and adenylate cyclase, abbreviated AC) is an enzyme with key regulatory roles in essentially all cells. It is the most polyphyletic known enzyme: six distinct classes have been described, all catalyzing the same reaction but representing unrelated gene families with no known sequence or structural homology. The best known class of adenylyl cyclases is class III or AC-III (Roman numerals are used for classes). AC-III occurs widely in eukaryotes and has important roles in many human tissues.
Contents
- Camp and adenylyl cyclase
- Adenylyl cyclase dependent pathway fast response
- Class I
- Class II
- Class III
- Structure
- Types
- Regulation
- Class IV
- Classes V and VI
- References
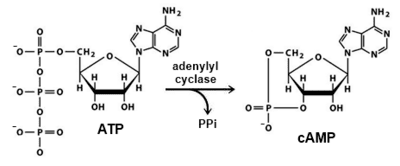
All classes of adenylyl cyclases catalyse the conversion of adenosine triphosphate (ATP) to 3',5'-cyclic AMP (cAMP) and pyrophosphate. Magnesium ions are generally required and appears to be closely involved in the enzymatic mechanism. The cAMP produced by AC then serves as a regulatory signal via specific cAMP-binding proteins, either transcription factors, enzymes (e.g., cAMP-dependent kinases), or ion transporters.
Adenylyl cyclase dependent pathway fast response
Class I

The first class of adenylyl cyclases occur in many bacteria including E. coli. This was the first class of AC to be characterized. It was observed that E. coli deprived of glucose produce cAMP that serves as an internal signal to activate expression of genes for importing and metabolizing other sugars. cAMP exerts this effect by binding the transcription factor CRP, also known as CAP. Class I AC's are large cytosolic enzymes (~100 kDa) with a large regulatory domain (~50 kDa) that indirectly senses glucose levels. As of 2012, no crystal structure is available for class I AC.
Class II
These adenylyl cyclases are toxins secreted by pathogenic bacteria such as Bacillus anthracis and Bordetella pertussis during infection. These bacteria also secrete proteins that enable the AC-II to enter host cells, where the exogenous AC activity undermines normal cellular processes. The genes for Class II AC's are known as cyaA. Several crystal structures are known for AC-II enzymes.
Class III
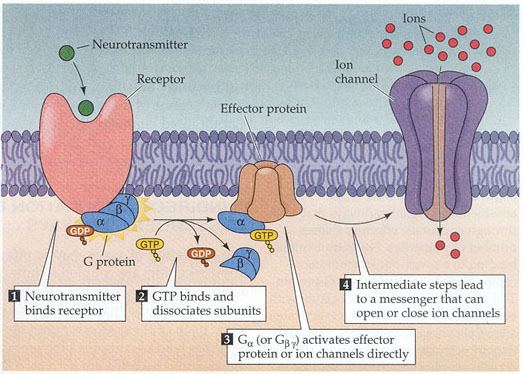
These adenylyl cyclases are the most familiar based on extensive study due to their important roles in human health. They are also found in some bacteria, notably Mycobacterium tuberculosis where they appear to have a key role in pathogenesis. Most AC-III's are integral membrane proteins involved in transducing extracellular signals into intracellular responses. A Nobel Prize was awarded to Earl Sutherland in 1971 for discovering the key role of AC-III in human liver, where adrenaline indirectly stimulates AC to mobilize stored energy in the "fight or flight" response. The effect of adrenaline is via a G protein signaling cascade, which transmits chemical signals from outside the cell across the membrane to the inside of the cell (cytoplasm). The outside signal (in this case, adrenaline) binds to a receptor, which transmits a signal to the G protein, which transmits a signal to adenylyl cyclase, which transmits a signal by converting adenosine triphosphate to cyclic adenosine monophosphate (cAMP). cAMP is known as a second messenger.
Cyclic AMP is an important molecule in eukaryotic signal transduction, a so-called second messenger. Adenylyl cyclases are often activated or inhibited by G proteins, which are coupled to membrane receptors and thus can respond to hormonal or other stimuli. Following activation of adenylyl cyclase, the resulting cAMP acts as a second messenger by interacting with and regulating other proteins such as protein kinase A and cyclic nucleotide-gated ion channels.
Photoactivatable adenylyl cyclase (PAC) was discovered in E. gracilis and can be expressed in other organisms through genetic manipulation. Shining blue light on a cell containing PAC activates it and abruptly increases the rate of conversion of ATP to cAMP. This is a useful technique for researchers in neuroscience because it allows them to quickly increase the intracellular cAMP levels in particular neurons, and to study the effect of that increase in neural activity on the behavior of the organism. For example, PAC expression in certain neurons has been shown to alter the grooming behavior in fruit flies exposed to blue light. Channelrhodopsin-2 is also used in a similar fashion.
Structure
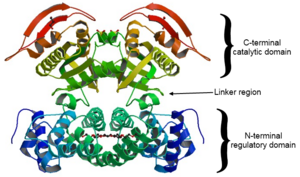
Most class III adenylyl cyclases are transmembrane proteins with 12 transmembrane segments. The protein is organized with 6 transmembrane segments, then the C1 cytoplasmic domain, then another 6 membrane segments, and then a second cytoplasmic domain called C2. The important parts for function are the N-terminus and the C1 and C2 regions. The C1a and C2a subdomains are homologous and form an intramolecular 'dimer' that forms the active site. In Mycobacterium tuberculosis, the AC-III polypeptide is only half as long, comprising one 6-transmembrane domain followed by a cytoplasmic domain, but two of these form a functional homodimer that resembles the mammalian architecture.
Types
There are ten known isoforms of adenylyl cyclases in mammals:
These are also sometimes called simply AC1, AC2, etc., and, somewhat confusingly, sometimes Roman numerals are used for these isoforms that all belong to the overall AC class III. They differ mainly in how they are regulated, and are differentially expressed in various tissues throughout mammalian development.
Regulation
Adenylyl cyclase is dually regulated by G proteins (Gs stimulating activity and Gi inhibiting it), and by forskolin, as well as other isoform-specific effectors:
In neurons, calcium-sensitive adenylyl cyclases are located next to calcium ion channels for faster reaction to Ca2+ influx; they are suspected of playing an important role in learning processes. This is supported by the fact that adenylyl cyclases are coincidence detectors, meaning that they are activated only by several different signals occurring together (source?). In peripheral cells and tissues adenylyl cyclases appear to form molecular complexes with specific receptors and other signaling proteins in an isoform-specific manner.
Class IV
AC-IV was first reported in the bacterium Aeromonas hydrophila, and the structure of the AC-IV from Yersinia pestis has been reported. These are the smallest of the AC enzyme classes; the AC-IV from Yersinia is a dimer of 19 kDa subunits with no known regulatory components.
Classes V and VI
These forms of AC have been reported in specific bacteria (Prevotella ruminicola and Rhizobium etti, respectively) and have not been extensively characterized.