
 | ||
The spine apparatus (SA) is a specialized form of endoplasmic reticulum (ER) that is found in a subpopulation of dendritic spines in central neurons. It was discovered by E. G. Gray in 1959 when he applied electron microscopy to fixed cortical tissue. The SA consists of a series of stacked discs that are thought to be connected to each other and to the dendritic system of ER-tubules. The actin binding protein synaptopodin (which was originally described in podocytes of the kidney) is an essential component of the SA. Mice that lack the gene for synaptopodin do not form a spine apparatus. The SA is believed to play a critical role in learning and memory, but the exact function of the spine apparatus is still considered relatively enigmatic.
Contents
Morphology
The spine apparatus consists of membranous saccules and tubules along with wispy filamentous material and is usually associated with a large mushroom-shaped dendritic spine. The wispy filamentous material is a cytoskeletal network which is responsible for the maintenance and alteration of spine shape and controls effectiveness of axospinous synapses. The morphology of the spine apparatus is highly indicative of and similar to the morphology and structure of the smooth surfaced endoplasmic reticulum of the dendrite. Consisting of continuous parallel flattened cisternae, the spine apparatus has a large surface area which is beneficial for its function.
Benefits of the large surface area of the spine apparatus include increased electronic properties of the spine and contribution to longitudinal resistance of the cytoplasm. The spine apparatus occupies a large portion of the volume of the spine stalk, which allows it to contribute significantly to the longitudinal resistance of the cytoplasm. Therefore, the spine apparatus can have a direct effect on the membrane potential of the spine plasma membrane by variation in position and volume.
The spine apparatus structure allows for dynamic changes in the surface area of the spine plasma membrane. For example, calcium-dependent mechanisms, similar to ones associated with cell shape and maintenance, have been linked to dynamic changes of spine plasma membrane surface area. These calcium-dependent mechanisms have a direct correlation with dynamic changes in dendritic spines, and hence spine plasma membrane surface area. The connection to the smooth endoplasmic reticulum also suggests a potential pathway for the transfer of proteins between the spine and dendrite. Also, the similarity between the smooth endoplasmic reticulum and the spine apparatus suggests that the spine apparatus could function as a reservoir for calcium ions.
The structures of spine apparatuses differ depending on their elaboration and position in the spine. Spines can be classified into four different categories: thin, stubby, mushroom, and branched. The shape of the spine apparatus differs depending upon which spine type it is associated with. For example, when the spine apparatus is associated with thin spines, the morphology is simple and consists of a very basic tubular form; however, when the spine apparatus is associated with mushroom shaped dendritic spines, the morphology is a complex laminar arrangement of saccules in the spine head and stalk.
Local Protein Synthesis and Trafficking
For some time, the function of the spine apparatus has been considered enigmatic. Recent evidence, however, suggests the spine apparatus may possess several distinct functions. After elucidating the structure of the spine apparatus, Spacek and Harris noted a continuation of the smooth endoplasmic reticulum into the spine apparatus, where it then takes on a lamellar structure. This observation suggests the SA might play a role in vesicular transport, although a specific mechanism is not yet clear.
Furthermore, Pierce et al. proposed that the spine apparatus may be involved in post-translational protein processing, similar to that observed in the golgi apparatus, and function in the post-translational processing of GluR1 and GluR2 subunits, which are locally translated in dentritic spines, of AMPA receptors. The spine apparatus has also been shown to be involved in the post-translational processing and spatial delivery of NMDA receptors, which also function as glutamate receptors and play a significant role in controlling synaptic plasticity. Considering immunostaining studies have identified NMDARs and AMPARs in the spine apparatus, it has been proposed that the spine apparatus may be critical to the localization of AMPARs and NMDARs to synapses during LTP formation.
The appearance of molecular markers for satellite secretory pathways provides further evidence that the spine apparatus plays a role in local integral membrane protein translocation and processing. More specifically, the protein translocation site marker (Sec61α) and the Golgi cisternae markers (giantin and α-mannosidase II) have been observed in the spine apparatus.
Calcium Signaling
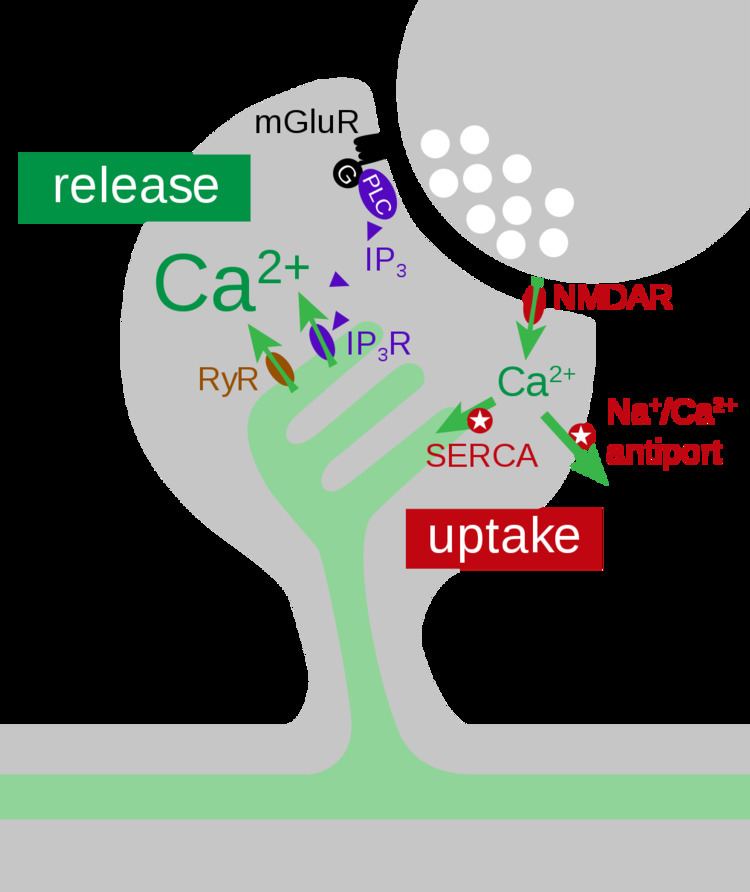
Synaptic activity triggers Ca2+ influx into dendritic spines via NMDA receptors and voltage-dependent calcium channels. Free Ca2+ ions are rapidly removed from the cytoplasm through Na+/Ca2+ exchangers in the plasma membrane and by sarco/endoplasmic reticulum Ca2+ ATPases (SERCA pumps) that mediate Ca2+ uptake into the smooth endoplasmic reticulum (sER). The spine apparatus, as a sub-compartment of the sER, has a large surface area and is thought to act as an efficient calcium buffer inside the spine (Figure 2).
Recent studies have shown that the spine apparatus is also able to release Ca2+ through inositol trisphosphate receptors (IP3Rs) or ryanodine receptors (RyRs). The calcium-sensitive nature of IP3Rs and RyRs makes both receptors capable of regenerative calcium-induced calcium release (CICR). In dendritic shafts and spines of hippocampal neurons the presence of both RyRs and IP3Rs has been shown by immunostaining. Ca2+ release is triggered by glutamate release, activating Group I metabotropic glutamate receptors (mGluRs). The downstream signaling cascade leads to elevated IP3 levels inside the spine (Figure 2) which trigger Ca2+ release events only in those spines that contain sER.
Plasticity
The spine apparatus’ ability to release calcium into the cytosol is thought to contribute to the development of synaptic plasticity. This was first shown in an experiment using synaptopodin(SP)-deficient mice, which do not show a spine apparatus in dendritic spines. These SP-deficient mice showed a decrease in long-term potentiation (LTP). Furthermore, LTP1 (short-term plasticity which requires post-translational protein modifications, but is protein synthesis independent), LTP2 (slowly decaying plasticity which is protein synthesis-dependent but does not require gene transcription modifications), and LTP3 (long-lasting LTP that is translation- and transcription-dependent) were decreased as a result of the absence of the spine apparatus.
Further studies have shown that altered expression and distribution of RyRs, IP3Rs, and L-type voltage-dependent calcium channels (L-VDCCs) can decrease LTP1, LTP2, and LTP3. This observation, combined with the observation that SP-deficient mice have decreased LTP, suggests the proper expression and distribution of calcium channels within the spine apparatus is required for controlling synaptic plasticity. Moreover, the spine apparatus is critical for maintaining the cytosolic calcium levels which are central to the formation of synaptic plasticity.
SP-deficient mice also exhibited behavioral changes, including decreased horizontal locomotor activity, decreased anxiety, and a decreased ability to acquire LTP3-related spatial memory, as shown in a radial arm maze. The decreased locomotor activity and decreased spatial learning suggest the spine apparatus’ role in inducing cerebellar LTP, in addition to hippocampal LTP. The exact mechanisms of these deficiencies are not fully understood. These behavioral modifications are thought to be epiphenomena of the absence of a spine apparatus and the changes in cytosolic calcium control mechanisms typically provided by the spine apparatus.
Synaptopodin, a protein that binds actin and α-actinin-2, is closely associated with the spine apparatus. Although SP-deficient mature mice both lack the spine apparatus and display impaired LTP, the relationship between SP, the spine apparatus, and plasticity is complicated by the following three findings: (1) In addition to being associated with the spine apparatus, SP is also located in the cisternal organelle, which is structurally similar to the spine apparatus. (2) The spine apparatus is generally found in mushroom spines of mature neurons, but ample levels of SP and expression of LTP have been detected in juvenile rats just 15 days old. (3) Unlike SP, the spine apparatus has not been found in cultured neurons. These findings have led some to the conclusion that the spine apparatus is implicated in plasticity only because this organelle is associated with SP.
Direct evidence for an essential function of the spine apparatus in mGluR-dependent long-term depression (LTD) has been provided by comparing plasticity of synapses on spines with or without spine apparatus. In this study, only synapses associated with sER displayed this form of depression. Blocking IP3-mediated Ca2+ release blocked synaptic depression, indicating a causal role of the spine apparatus in the induction of mGluR-dependent LTD.
Relevance to Disease
Recent evidence suggests that structural changes in the spine apparatus may be linked to brain disorders. When parkinsonian symptoms were induced in monkeys via treatment with MPTP, alterations in morphology were observed, including an increase in the ratio of spine apparatus volume to spine volume. Abnormal spine apparatus morphology has also been noted in peritumorous and edematous tissues of the human brain, and these changes in structure may result in compromised or altered function.
Much has yet to be discovered about the role of the spine apparatus in brain disease; the specific effects of changes in SA morphology remain a topic of study. Scientists are hopeful that serial section electron microscopy (ssEM), a high resolution technique that can be used to visualize dendritic spines and their intracellular components, will allow for a greater understanding of the relationship between alterations in SA structure and disease pathology.
Alterations to the morphology of the spine apparatus have also been observed in rats exposed to chronic levels of ethanol. Atrophied spine apparatuses were observed in these animals, as well as in anesthetized animals. The exact mechanisms involved in the size reduction and atrophy of the spine apparatus are unknown, but it is thought that these mechanisms may contribute to changes in LTP and behavior.