
Group Group IV ((+)ssRNA) Scientific name Picornaviridae | Rank Family | |
 | ||
Lower classifications | ||
Picornavirus overview video sketchymicro usmle microbiology review
A picornavirus is a virus belonging to the family Picornaviridae, a family of viruses in the order Picornavirales. Vertebrates, including humans, serve as natural hosts. Picornaviruses are nonenveloped viruses that represent a large family of small, cytoplasmic, plus-strand RNA(~7.5kb) viruses with a 30 nm icosahedral capsid. Its genome does not have a lipid membrane. Picornaviruses are found in mammals and birds. There are currently 50 species in this family, divided among 29 genera. These include the Entero-, Aphtho-, Cardio-, Rhino- and Hepatovirus genera. The viruses in this family can cause a range of diseases including paralysis, meningitis, hepatitis and poliomyelitis. Picornaviruses are in Baltimore IV class. Their genome single-stranded (+) sense RNA is that functions as mRNA after entry into the cell and all viral mRNA synthesized is of genome polarity. The mRNA encodes RNA dependent RNA polymerase. This polymerase makes complementary minus strands of RNA, then uses them as templates to make more plus strands. The first synthesized molecule is protein in contrast to all other viruses. So, Overview of picornavirus replication are in order to attachment, entry, translation, transcription/genome replication (one and the same process), assembly and exit.
Contents
Virology lecture picornaviruses
Diseases
Enteroviruses infect the enteric tract, which is reflected in their name. On the other hand, rhinoviruses infect primarily the nose and the throat. Enteroviruses replicate at 37 °C, whereas rhinoviruses grow better at 33 °C, as this is the lower temperature of the nose. Enteroviruses are stable under acid conditions and thus they are able to survive exposure to gastric acid. In contrast, rhinoviruses are acid-labile (inactivated or destroyed by low pH conditions) and that is the reason why rhinovirus infections are restricted to the nose and throat.
Taxonomy
Group: ssRNA(+)
Structure
Picornaviruses are non-enveloped, with an icosahedral capsid. The capsid is an arrangement of 60 protomers in a tightly packed icosahedral structure. Each protomer consists of 4 polypeptides known as VP (viral protein) 1, 2, 3 and 4. VP2 and VP4 polypeptides originate from one protomer known as VP0 that is cleaved to give the different capsid components. The icosahedral is said to have a triangulation number of 3, this means that in the icosahedral structure each of the 60 triangles that make up the capsid are split into 3 little triangles with a subunit on the corner.In many picornaviruses have a deep cleft formed by around each of the 12 vertices of icosahedrons.The outer surface of the capsid is composed of regions of VP1,VP2 and VP3. Around each of the vertices is a canyon lined with the C termini of VP1 and VP3. The interior surface of the capsid is composed of VP4 and the N termini of VP1. J.Esposito and Professor Freederick A. Murphy demonstrates cleft structure referred to as canyons, using X-ray crystallography and cryo-electron microscopy. Depending on the type and degree of dehydration the viral particle is around 27–30 nm in diameter. The viral genome is around 2500 nm in length so we can therefore conclude that it must be tightly packaged within the capsid along with substances such as sodium ions in order to cancel out the negative charges on the RNA caused by the phosphate groups.
Genome
Picornaviruses are classed under Baltimore's viral classification system as group IV viruses as they contain a single stranded, positive sense RNA genome. Their genome ranges between 7.1 and 8.9 kb (kilobases) in length. Like most positive sense RNA genomes, the genetic material alone is infectious; although substantially less virulent than if contained within the viral particle, the RNA can have increased infectivity when transfected into cells. The genome RNA is unusual because it has a protein on the 5' end that is used as a primer for transcription by RNA polymerase.This primer is called VPg genome range between 2–3 kb. VPg contain tyrosine residue at the 3’ end. Tyrosine as a –OH source for covalently linked to 5’ end of RNA.
The genome is non-segmented and positive-sense (the same sense as mammalian mRNA, being read 5' to 3'). Unlike mammalian mRNA picornaviruses do not have a 5' cap but a virally encoded protein known as VPg. However, like mammalian mRNA, the genome does have a poly(A) tail at the 3' end. There is an un-translated region (UTR) at both ends of the picornavirus genome. The 5' UTR is usually longer, being around 500–1200 nucleotides (nt) in length, compared to that of the 3' UTR, which is around 30–650 nt. It is thought that the 5' UTR is important in translation and the 3' in negative strand synthesis; however the 5' end may also have a role to play in virulence of the virus. The rest of the genome encodes structural proteins at the 5' end and non-structural proteins at the 3' end in a single polyprotein.
The polyprotein is organised as follows: L-1ABCD-2ABC-3ABCD with each letter representing a protein, however, there are variations to this layout.
The 1A, 1B, 1C, and 1D proteins are the capsid proteins VP4, VP2, VP3, and VP1, respectively.Virus-coded proteases perform the cleavages, some of which are intramolecular. The polyprotein is first cut to yield P1, P2 and P3. P1 becomes myristylated at the N terminus before being cleaved to VP0, VP3 and VP1, the proteins that will form procapsids; VP0 will later be cleaved to produce VP2 and VP4. Other cleavage products include 3B (VPg), 2C (an ATPase) and 3D (the RNA polymerase).
Replication
Genomic RNAs of picornaviruses possess multiple RNA elements and they are required for both negative and plus strand RNA synthesis. The cis acting replication(cre) element is a required for replication. The stem-loop-structure contains the cre is independent of position but it changes with location between virus types when it has been identified. Also, the 3’ end elements of viral RNA are significant and efficient for RNA replication of picornaviruses. The 3’ end of picornavirus contains poly(A) tract which be required for infectivity. On the other hand, RNA synthesis is hypothesized to occur in this region.3’ end NCR of poliovirus is not necessary for negative-strands synthesis. However, it is important element for positive—strand synthesis. Additionally,5’ end NCR that contain secondary structural elements is required for RNA replication and poliovirus translation initiation(IRES). Internal Ribosome Entry Site (IRES) are RNA structures that allow cap independent initiation of translation, and are able to initiate translation in the middle of a messenger RNA.
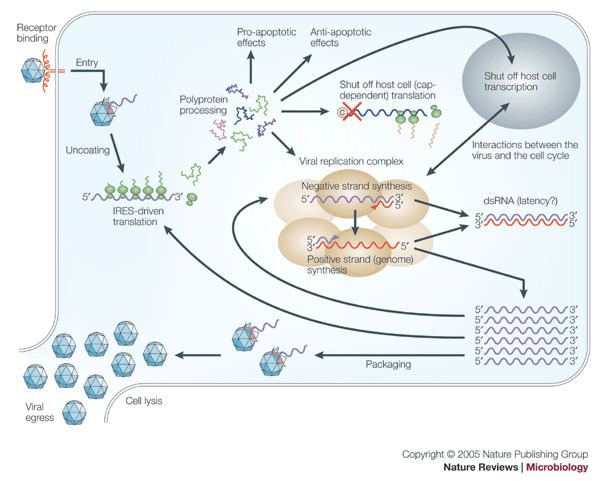
The viral particle binds to cell surface receptors. Cell surface receptors are characterized for each serotype of picornaviruses. For example, poliovirus receptor is glycoprotein CD155 which is special receptor for human and some other primate species. For this reason, poliovirus couldn’t be made in many laboratories until transgenic mice having a CD155 receptor on their cell surface were developed in the 1990s. These animals can be infected and used for studies of replication and pathogenesis. Binding causes a conformational change in the viral capsid proteins, and myristic acid are released. These acids form a pore in the cell membrane through which RNA is injected [2]. Once inside the cell, the RNA un-coats and the (+) strand RNA genome is replicated through a double-stranded RNA intermediate that is formed using viral RDRP (RNA-Dependent RNA polymerase). Translation by host cell ribosomes is not initiated by a 5' G cap as usual, but rather is initiated by an IRES (Internal Ribosome Entry Site). The viral lifecycle is very rapid with the whole process of replication being completed on average within 8 hours. However, as little as 30 minutes after initial infection, cell protein synthesis declines to almost zero output – essentially the macromolecular synthesis of cell proteins is shut off. Over the next 1–2 hours there is a loss of margination of chromatin and homogeneity in the nucleus, before the viral proteins start to be synthesized and a vacuole appears in the cytoplasm close to the nucleus that gradually starts to spread as the time after infection reaches around 3 hours. After this time the cell plasma membrane becomes permeable, at 4–6 hours the virus particles assemble, and can sometimes be seen in the cytoplasm. At around 8 hours the cell is effectively dead and lyses to release the viral particles.
Experimental data from single step growth-curve-like experiments have allowed scientists to look at the replication of the picornaviruses in great detail. The whole of replication occurs within the host cell cytoplasm and infection can even happen in cells that do not contain a nucleus (known as enucleated cells) and those treated with actinomycin D (this antibiotic would inhibit viral replication if this occurred in the nucleus.)
Translation takes place by -1 ribosomal frameshifting, viral initiation, and ribosomal skipping. The virus exits the host cell by lysis, and viroporins. Vertebrates serve as the natural host. Transmission routes are fecal-oral, contact, ingestion, and air borne particles.
VPg
Picornaviruses have a viral protein (VPg) covalently linked to 5’ end of their genomes instead of 7-methylguanosine cap like cellular mRNAs. Virus RNA polymerases use VPg as primer. VPg as primer uses both minus and plus strand RNA synthesis. Picornavirus replication is initiated by the uridylylation of viral protein genome-linked (VPg).It is uridylylated at the hydroxyl group of a tyrosine residue. A VPg primer mechanism is utilized by the picornavirus (entero- aphtho- and others), additional virus groups (poty-, como-, calici- and others) and picornavirus-like (coronavirus, notavirus, etc.) supergroup of RNA viruses. The mechanism has been best studied for the enteroviruses (which include many human pathogens, such as poliovirus and coxsackie viruses) as well as for the aphthovirus, an animal pathogen causing foot and mouth disease (FMDV).
In this group, primer-dependent RNA synthesis utilizes a small 22–25 amino acid long viral protein linked to the genome (VPg) to initiate polymerase activity, where the primer is covalently bound to the 5’ end of the RNA template. The uridylylation occurs at a tyrosine residue at the third position of the VPg. A cis-acting replication element (CRE), which is a RNA stem loop structure, serves as a template for the uridylylation of VPg, resulting in the synthesis of VPgpUpUOH. Mutations within the CRE-RNA structure prevent VPg uridylylation, and mutations within the VPg sequence can severely diminish RdRp catalytic activity. While the tyrosine hydroxyl of VPg can prime negative-strand RNA synthesis in a CRE- and VPgpUpUOH-independent manner, CRE-dependent VPgpUpUOH synthesis is absolutely required for positive-strand RNA synthesis. CRE-dependent VPg uridylylation lowers the Km¬ of UTP required for viral RNA replication and CRE-dependent VPgpUpUOH synthesis, and is required for efficient negative-strand RNA synthesis, especially when UTP concentrations are limiting. The VPgpUpUOH primer is transferred to the 3’ end of the RNA template for elongation, which can continue by addition of nucleotide bases by RdRp. Partial crystal structures for VPgs of foot and mouth disease virus and coxsackie virus B3 suggest that there may be two sites on the viral polymerase for the small VPgs of the picornaviruses. NMR solution structures of poliovirus VPg and VPgpU show that uridylylation stabilizes the structure of the VPg, which is otherwise quite flexible in solution. The second site may be used for uridylylation, after which the VPgpU can initiate RNA synthesis. It should be noted that the VPg primers of caliciviruses, whose structures are only beginning to be revealed, are much larger than those of the picornaviruses. Mechanisms for uridylylation and priming may be quite different in all of these groups.
VPg uridylylation may include the use of precursor proteins, allowing for the determination of a possible mechanism for the location of the diuridylylated, VPg-containing precursor at the 3’ end of plus- or minus-strand RNA for production of full-length RNA. Determinants of VPg uridylylation efficiency suggest formation and/or collapse or release of the uridylylated product as the rate-limiting step in vitro depending upon the VPg donor employed. Precursor proteins also have an effect on VPg-CRE specificity and stability. The upper RNA stem loop, to which VPg binds, has a significant impact on both retention, and recruitment, of VPg and Pol. The stem loop of CRE will partially unwind, allowing the precursor components to bind and recruit VPg and Pol4. The CRE loop has a defined consensus sequence to which the initiation components bind, however; there is no consensus sequence for the supporting stem, which suggests that only the structural stability of the CRE is important.
Assembly and organization of the picornavirus VPg ribonucleoprotein complex.
VPg may also play an important role in specific recognition of viral genome by movement protein (MP). Movement proteins are non-structural proteins encoded by many, if not all, plant viruses to enable their movement from one infected cell to neighboring cells. MP and VPg interact to provide specificity for the transport of viral RNA from cell to cell. To fulfill energy requirements, MP also interacts with P10, which is a cellular ATPase.
History
In 1897, foot-and-mouth disease virus (FMDV), the first animal virus, was discovered. FMDV is the prototypic member of the Aphthovirus genus in the Picornaviridae family. The plaque assay was developed using poliovirus; the discovery of viral replication in culture was also with poliovirus in 1949. This was the first time that infection virus had been produced from molecular building blocks in the cells. Polyprotein synthesis, internal ribosome entry sites, and uncapped mRNA were all discovered by studying poliovirus infected cells, and a poliovirus clone was the first infectious DNA clone made of an RNA virus in animals. Along with rhinovirus, poliovirus was the first animal virus to have its structure determined by x-ray crystallography. RNA dependent RNA polymerase was discovered in Mengovirus, a genus of picornaviruses.
Related Families
Within the order Picornavirales, there are related viral families, such as the plant infecting Secoviridae, and the insect infecting Dicistroviridae.
Secoviridae
The plant-infecting picorna-like viruses have a number of properties that are distinct from the animal viruses.The picornaviruses include the plant-infecting Secovirida. It has both icosahedral virus particles, viral RNA-dependent RNA polymerase and protease and viral replication proteins. But they have distinguished properties at the same time. For example, secoviruses infect plants and it has specialized proteins. The effect of secoviruses is important on cultivated crops. It infected a wide range of plants from grapevine to rice. They have been classified into the family Secoviridae containing the subfamily Comovirinae (genera Comovirus, Fabavirus and Nepovirus), and genera Sequivirus, Waikavirus, Cheravirus, Sadwavirus, and Torradovirus (type species Tomato torrado virus)).
Dicistroviridae
Members of the family dicistroviruses are to beneficial invertebrates such as honey bees and shrimp and to insect pests of medical and agricultural importance that share features with animal and human viruses of the family. Picornaviridae and other insect or marine viruses of the order Picornavirales. Its genome is linear, single stranded positive sense RNA with a viral genome-linked protein (VPg) covalently linked at the 5’ end and a 3’ poly (A) tract as picornaviruses. Plautia stali intestine virus kelp fly virus, Ectropis obliqua picorna-like virus, deformed wing virus, acute bee paralysis virus, Drosophila C virus, Rhopalosiphum padi virus, and Himetobi P virus. Several have been placed in a separate family—the Dicistroviridae. Others have been placed into a new family Iflaviridae. This family includes Infectious flacherie virus and SeIV-1 virus. Another virus is Nora virus from Drosophila melanogaster. This latter virus awaits further classification.