
 | ||
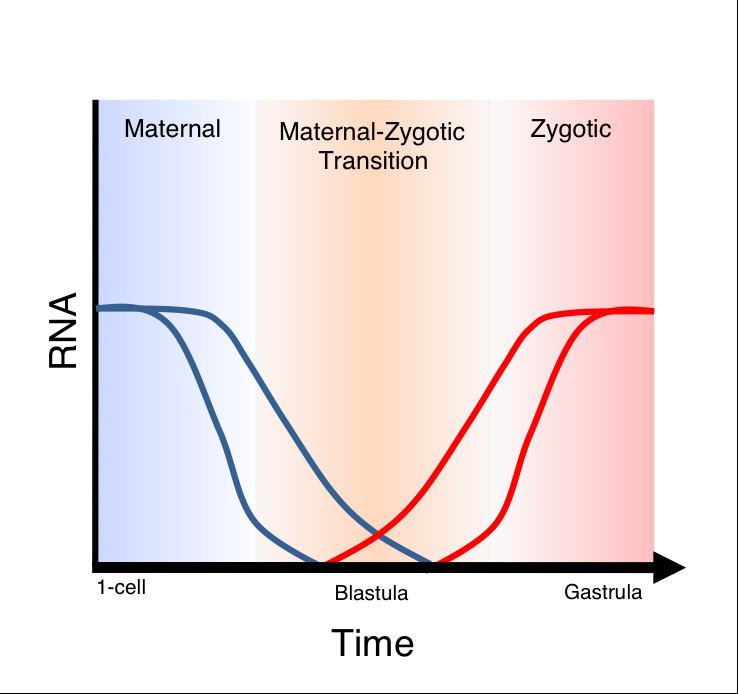
Maternal to zygotic transition (MZT) is the stage in embryonic development during which development comes under the exclusive control of the zygotic genome. This requires both zygotic genome activation (ZGA) and degradation of maternal products. MZT is often thought to be synonymous with midblastula transition (MBT), but these processes are, in fact, distinct. However, the MBT roughly coincides with ZGA in many metazoans, and thus may share some common regulatory features. For example, both processes are proposed to be regulated by the nucleocytoplasmic ratio. MBT strictly refers to changes in the cell cycle and cell motility that occur just prior to gastrulation. In the early cleavage stages of embryogenesis, rapid divisions occur synchronously and there are no "gap" stages in the cell cycle. During these stages, there is also little to no transcription of mRNA from the zygotic genome, but zygotic transcription is not required for MBT to occur. Cellular functions during early cleavage are carried out primarily by maternal products – proteins and mRNAs contributed to the egg during oogenesis.
Contents
Zygotic genome activation
To begin transcription of zygotic genes, the embryo must first overcome the silencing that has been established. The cause of this silencing could be due to several factors: chromatin modifications leading to repression, lack of adequate transcription machinery, or lack of time in which significant transcription can occur due to the shortened cell cycles. Evidence for the first method was provided by Newport and Kirschner's experiments showing that nucleocytoplasmic ratio plays a role in activating zygotic transcription. They suggest that a defined amount of repressor is packaged into the egg, and that the exponential amplification of DNA at each cell cycle results in titration of the repressor at the appropriate time. Indeed, in Xenopus embryos in which excess DNA is introduced, transcription begins earlier. More recently, evidence has been shown that transcription of a subset of genes in Drosophila is delayed by one cell cycle in haploid embryos. The second mechanism of repression has also been addressed experimentally. Prioleau et al. show that by introducing TATA binding protein (TBP) into Xenopus oocytes, the block in transcription can be partially overcome. The hypothesis that shortened cell cycles can cause repression of transcription is supported by the observation that mitosis causes transcription to cease.
Clearing of maternal transcripts
To eliminate the contribution of maternal gene products to development, maternally-supplied mRNAs must be degraded in the embryo. Studies in Drosophila have shown that sequences in the 3' UTR of maternal transcripts mediate their degradation (reviewed by Tadros and Lipshitz) These sequences are recognized by regulatory proteins that cause destabilization or degradation of the transcripts. Recent studies in both zebrafish and Xenopus have found evidence of a role for microRNAs in degradation of maternal transcripts. In zebrafish, the microRNA miR-430 is expressed at the onset of zygotic transcription and targets several hundred mRNAs for deadenylation and degradation. Many of these targets are genes that are expressed maternally. Similarly, in Xenopus, the miR-430 ortholog miR-427 has been shown to target maternal mRNAs for deadenylation. Specifically, miR-427 targets include cell cycle regulators such as Cyclin A1 and Cyclin B2.