
 | ||
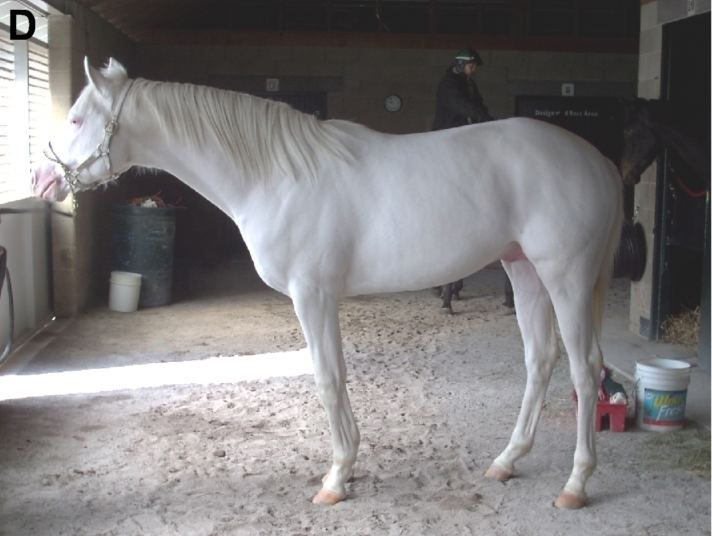
Dominant white is a group of genetically related coat color conditions in the horse, best known for producing an all-white coat, but also for producing some forms of white spotting and white markings. Dominant white horses are born with unpigmented pink skin and white hair with dark eyes, although the amount of white hair or spotting can vary depending on which genetic mutation is involved. Dominant white is a rare condition, and under normal conditions, at least one parent must be dominant white to produce dominant white offspring. However, most of the currently-known alleles of dominant white can be linked to a documented spontaneous mutation in a single ancestor.
Contents
- Identification
- Prevalence
- Inheritance
- History of dominant white research
- Allelic series
- Molecular genetics
- Lethality
- White horses that are not dominant white
- Non white colors
- Lethal White
- Sabino
- Mosaicism
- Homologous conditions
- References
Dominant white can occur in any breed, and has been studied in many different breeds. Two color breeds, the American White Horse and Camarillo White Horse are characterized by their dominant white coats.
There are many different forms of dominant white; in genetics, as of 2013 they are labeled W1 through W20. All known dominant white coat colors are associated with the KIT gene. As the name suggests, these known white coats are inherited dominantly, meaning that a horse only needs one copy of a W allele to have the white or white spotted coat.
Dominant white is genetically distinct from Sabino and both genetically and visually distinct from gray and cremello. Dominant white is not the same as lethal white syndrome, nor are dominant white horses "albinos". Albinism has never been documented in horses. Some forms of dominant white are thought to result in nonviable embryos when a zygote has two W alleles (is homozygous). However, this has not been verified for all dominant white genetic variations.
Identification
Dominant white horses are born with pink skin and a white coat, which they retain throughout their lives. Although the term "dominant white" is typically associated with a pure white coat, such horses may be all-white, near-white, partially white, or exhibit an irregular spotting pattern similar to that of sabino horses. The amount of white hair depends on which mutation of W is involved. Non-white areas of skin and hair are most commonly seen along the dorsal midline of the horse, known as the topline, and are especially common in the mane and on the ears. They may also have interspersed specks or spots of non-white skin and hair. In addition, the hooves are most often white, but may have striping if there is pigmented skin on the coronary band just above the hoof. In some cases, foals born with residual non-white hair may lose some or all of this pigment with age, without the help of the gray factor. Dominant white spotting does not affect eye color, and most dominant white horses have brown eyes. The pink skin is devoid of pigment cells (melanocytes), and appears pink from the underlying network of capillaries. White hair is rooted in unpigmented pink skin. There are many other genetic factors that produce white, near-white, and off-white coat colors in horses, some of which are visually very similar to dominant white.
Prevalence
Dominant white is one of several potential genetic causes for horses with near-white or completely white coats; it may occur through spontaneous mutation, and thus may be found unexpectedly in any breed, even those that discourage excessive white markings. To date, dominant white has been identified in multiple families of Thoroughbreds, American Quarter Horses, Frederiksborg horses, Icelandic horses, Shetland ponies, Franches Montagnes horses, South German Draft horses, and in one family of the Arabian horse. The American White Horse, which is descended primarily from one dominant white stallion crossed on non-white mares, is known for its dominant white coat, as is the Camarillo White Horse.
Inheritance
The W locus was mapped to the KIT gene in 2007. The terms "Kit oncogene" and "dominant spotting" gene, symbolized by KIT and W respectively, can be used interchangeably. Current research has now shown that there are multiple forms, or alleles, of the W gene. All horses possess the KIT gene, as it is necessary for survival even at the earliest stages of development. The presence or absence of dominant white is based on the presence of certain altered forms of KIT. Each unique form is called an allele, and for every trait, all animals inherit one allele from each parent. The original or "normal" form of KIT, which is expected in horses without dominant white spotting, is called the "wild type" allele. Thus, a dominant white horse has one KIT allele with a mutation associated with dominant white spotting, and one wild type KIT allele.
History of dominant white research
Dominant white horses were first described in scientific literature in 1912. Horse breeder William P. Newell described his family of white and near-white horses to researcher A. P. Sturtevant of Columbia University:
"The colour of skin is white or so-called pink, usually with a few small dark specks in skin. Some have a great many dark spots in skin. These latter usually have a few dark stripes in hoofs; otherwise the hoofs are almost invariably white. Those that do not have dark specks in skin usually have glass or watch eyes, otherwise dark eyes ... I have one colt coming one year old that is pure white, not a coloured speck on him, not a coloured hair on him, and with glass [blue] eyes."
Sturtevant and his contemporaries agreed that this colt's blue eyes were inherited separately from his white coat. In 1912, Sturtevant assigned the "white" trait to the White or W locus. At the time there was no means of assigning W to a position on the chromosome, or to a gene.
This family of white horses produced Old King in 1908, a dark-eyed white stallion that was purchased by Caleb R. and Hudson B. Thompson. Old King was bred to Morgan mares to produce a breed of horse known today as the American White Horse. A grandson of Old King, Snow King, was at the center of the first major study of the dominant white coat color in horses, conducted in 1969 by Dr. William L. Pulos of Alfred University and Dr. Frederick B. Hutt of Cornell. They concluded, based on test matings and progeny phenotype ratios, that the white coat was dominantly inherited and embryonic lethal in the homozygous state. Other factors, such as variations in expressivity and the influence of multiple genes, may have influenced the progeny ratios that Pulos and Hutt observed. The white coat of the American White Horse has not yet been mapped.
A 1924 study by C. Wriedt identified a heritable white coat color in the Frederiksborg horse. Wriedt described a range of what he considered to be homozygote phenotypes: all-white, white with pigmented flecks, or weißgraue, which transliterates to "white-gray." The German term for gray horse is schimmel, not weißgraue. Heterozygotes, according to Wriedt, ranged from roaned or diluted to more or less solid white horses. Reviewers, such as Miguel Odriozola, reinterpreted Wriedt's data in successive years, while Pulos and Hutt felt that his work had been "erroneous" because Wriedt never concluded that white was lethal when homozygous.
Other researchers prior to modern DNA analysis developed remarkably prescient theories. The gene itself was first proposed and named W in 1948. In a 1969 work on horse coat colors, A los colores del caballo, Miguel Odriozola suggested that various forms of dominantly inherited white spotting might be arranged sequentially along one chromosome, thus allowing for the varied expression of dominant white. He also proposed that other, distant genes might also influence the amount of white present.
Between the time of Pulos and Hutt's study in 1969 and the beginning of molecular-level research into dominant white in the 21st century, a pattern known as "Sabino" became regarded by some as a more likely cause of white phenotypes. Sabino is a type of white spotting, and the one allele now mapped, the dominantly inherited Sabino-1 (SB-1), is genetically related, though distinct. When homozygous, SB-1 can produce nearly all-white horses that resemble dominant white. Other genes responsible for all possible patterns labeled "Sabino" have not yet been identified, though some forms of the splashed white gene may be responsible for certain patterns.
In 2007, researchers from Switzerland and the United States published a paper identifying the genetic cause of dominant white spotting in horses from the Franches Montagnes horse, Camarillo White Horse, Arabian horse and Thoroughbred breeds. Each of these dominant white conditions had occurred separately and spontaneously in the past 75 years, and each represents a different allele (variation or form) of the same gene. These same researchers identified a further seven unique causes of dominant white in 2009: three in distinct families of Thoroughbreds, one Icelandic horse, one Holsteiner, a large family of American Quarter Horses and a family of South German Draft horses.
Allelic series
The KIT gene contains over 2000 base pairs, and a change in any of those base pairs results in a mutant allele. Over forty such alleles have been identified by sequencing the KIT genes of various horses. The resultant phenotype of most of these alleles is not yet known, but 20 have been linked to dominant white. To date, DNA tests can identify if a horse carries the various identified W alleles, some commercially available.
These alleles do not account for all dominantly inherited white spotting in horses. More KIT alleles are expected to be found with roles in white spotting. Most W alleles each occur within a specific breed or family and arise as spontaneous mutations. The KIT gene itself seems prone to mutation, and so new alleles of W could occur in virtually any breed.
Molecular genetics
The KIT gene encodes a protein called steel factor receptor, which is critical to the differentiation of stem cells into blood cells, sperm cells, and pigment cells. A process called alternative splicing, which uses the information encoded in the KIT gene to make slightly different proteins (isoforms) for use in different circumstances, may impact whether a mutation on KIT affects blood cells, sperm cells, or pigment cells. Steel factor receptor interacts chemically with steel factor or stem cell factor to relay chemical messages. These messages are used during embryonic development to signal the migration of early melanocytes (pigment cells) from the neural crest tissue to their eventual destinations in the dermal layer. The neural crest is a transient tissue in the embryo that lies along the dorsal line. Melanocytes migrate along the dorsal line to a number of specific sites: near the eye, near the ear, and the top of the head; six sites along each side of the body, and a few along the tail. At these sites, the cells undergo a few rounds of replication and differentiation, and then migrate down and around the body from the dorsal aspect towards the ventral aspect and the limb buds.
The timing of this migration is critical; all white markings, from a small star to a pure white coat, are caused by the failed migration of melanocytes.
A certain degree of the eventual amount of white, and its "design", is completely random. The development of an organism from single-celled to fully formed is a process with many, many steps. Even beginning with identical genomes, as in clones and identical twins, the process is unlikely to occur the same way twice. A process with this element of randomness is called a stochastic process, and cell differentiation is, in part, a stochastic process. The stochastic element of development is partly responsible for the eventual appearance of white on a horse, potentially accounting for nearly a quarter of the phenotype. The research team that studied dominant white cited "subtle variations in the amount of residual KIT protein" as a potential cause for the variability in phenotype of horses with the same kind of dominant white.
Lethality
Early embryonal lethality, also known as early embryonic death or a non-viable embryo, may occur when the embryo possesses two dominant white alleles, or have the homozygous genotype. The reason for this is that many mutations for W are caused by nonsense mutations, frameshift mutations or DNA deletions, which, if homozygous, would make it impossible to produce a functional KIT protein. However, it is possible that homozygous embryos from alleles of missense and splice site mutations might be viable because they have less effect on gene function. A 2013 study also unearthed horses that were compound W5/W20 heterozygotes, almost completely white, essentially with greater depigmentation than could be accounted for by either allele alone.
The embryonic lethality hypothesis was originally supported by Pulos and Hutt's 1969 study of Mendelian progeny ratios. Conclusions about Mendelian traits that are controlled by a single gene can be drawn from test breedings with large sample sizes. However, traits that are controlled by allelic series or multiple loci are not Mendelian characters, and are not subject to Mendelian ratios.
Pulos and Hutt knew that if the allele that created a white coat was recessive, then white horses would have to be homozygous for the condition and therefore breeding white horses together would always result in a white foal. However, this did not occur in their study and they concluded that white was not recessive. Conversely, if a white coat was a simple autosomal dominant, ww horses would be non-white, while both Ww and WW horses would be white, and the latter would always produce white offspring. But Pulos and Hutt did not observe any white horses that always produced white offspring, suggesting that homozygous dominant (WW) white horses did not exist. As a result, Pulos and Hutt concluded that white was semidominant and lethal in the homozygous state: ww horses were non-white, Ww were white, and WW died.
Pulos and Hutt reported that neonatal death rates in white foals were similar to those in non-white foals, and concluded that homozygous white fetuses died during gestation. No aborted fetuses were found, suggesting that death occurred early on in embryonic or fetal development and that the fetus was "resorbed."
Prior to Pulos and Hutt's work, researchers were split on the mode of inheritance of white and whether it was deleterious (harmful). Recent research has discovered several possible genetic pathways to a white coat, so disparities in these historical findings may reflect the action of different genes. It is also possible that the varied origins of Pulos and Hutt's white horses might be responsible for the lack of homozygotes. Therefore, it remains to be proven whether all equine dominant white mutations cause embryonic lethality in the homozygous state.
The white (W) locus was first recognized in mice in 1908. The mutation of the same name produces a belly spot and interspersed white hairs on the dorsal aspect of the coat in the heterozygote (W/+) and black-eyed white in the homozygote (W/W). While heterozygotes are healthy, homozygous W mice have severe macrocytic anemia and die within days. A mutation which affects multiple systems is "pleiotropic." Following the mapping of the KIT gene to the W locus in 1988, researchers began identifying other mutations as part of an allelic series of W. There are over 90 known W alleles, each representing a unique mutation on the KIT gene, which primarily produce white spotting from tiny head spots to fully white coats, macrocytic anemia from mild to lethal, and sterility. Some alleles, such as sash produce white spotting alone, while others affect the health of the animal even in the heterozygous state. Alleles encoding small amounts of white are no more likely to be linked with anemia and sterility than those encoding conspicuous white. Presently, no anecdotal or research evidence has suggested that equine KIT mutations affect health or fertility. A recent study showed that blood parameters in horses with the W1 mutation were normal.
"White" horses that are not dominant white
White horses are potent symbols in many cultures. An array of horse coat colors may be identified as "white," often inaccurately, and many are genetically distinct from "dominant white."
"Albino" horses have never been documented, despite references to so-called "albino" horses. Dominant white is caused by the absence of pigment cells (melanocytes), whereas albino animals have a normal distribution of melanocytes. Also, a diagnosis of albinism in humans is based on visual impairment, which has not been described in horses with dominant white nor similar coat colors. In other mammals, the diagnosis of albinism is based on the impairment of tyrosinase production through defects in the Color (C) gene. No mutations of the tyrosinase or C gene are known in horses.
Non-white colors
Lethal White
Foals with Lethal white syndrome (LWS) have two copies of the Frame overo gene and are born with white or nearly white coats and pink skin. However, unlike dominant white horses, foals with LWS are born with an underdeveloped colon that is untreatable, and if not euthanized, invariably die of colic within a few days of birth. Horses that carry only one allele of the LWS gene are healthy and typically exhibit the "frame overo" spotting pattern. In cases of "solid" horses with frame overo ancestry, uncertain "overo" (non-tobiano) phenotype, or horses with multiple patterns, the LWS allele can be detected by DNA test.
Sabino
Both dominant white and "Sabino-White" horses are identified by all-white or near-white coats with underlying pink skin and dark eyes, often with residual pigment along the dorsal midline. However, there are genetic differences. The term "dominant white" is reserved for known W alleles. Dominant white horses are heterozygous for any one of 11 known alleles of the KIT gene (e.g. W8/+). Homozygosity for some of the 11 known alleles may not create a viable embryo. In contrast, Sabino-White horses are homozygous for the Sabino 1 allele of the KIT gene (SB1/SB1).
Another type of sabino patterning, called simply "sabino," "minimal sabino" if slight, or if particularly dramatic, "maximum sabino," refers to horses that test negative for any of the Dominant White alleles, negative for Sabino 1, and also negative for Tobiano and Frame overo. Initially, dominant white was separated from sabino on the grounds that the former had to be entirely white, while the latter could possess some pigment. However, the 2007 and 2009 studies of dominant white showed that many dominant white alleles produce a range of white phenotypes that include horses with pigmented spots in hair and skin. Each of the larger families of dominant white studied included pure-white horses, horses described as having "sabino-like" white markings, as well as white horses described as "maximal sabino."
More recently, dominant white and sabino were distinguished from one another on the grounds that dominant white alleles produce nonviable embryos in the homozygous state, while Sabino 1 was viable when homozygous. However, not all KIT alleles currently identified as "dominant white" have been proven lethal.
The similarities between Dominant White, Sabino 1, and other forms of sabino may reflect their common molecular origin: The W1-W11 series and SB1 have been mapped to KIT. The researchers who mapped Sabino 1 suggested that other sabino-like patterns might also map to KIT. Similarly, major alleles for white leg and facial markings have also been mapped to or near to the KIT gene.
Mosaicism
Mosaicism in horses is thought to account for some spontaneous occurrences of white, near-white, spotted, and roan horses. Mosaicism refers to mutations that occur after the single-cell stage, and therefore affect only a portion of the adult cells. Mosaicism may be one possible cause for the rare occurrence of brindle coloring in horses. Mosaic-white horses would be visually indistinguishable from dominant whites. Mosaicism could produce white or partially white foals if a stem cell in the developing foal underwent a mutation, or change to the DNA, that resulted in unpigmented skin and hair. The cells that descend from the affected stem cell will exhibit the mutation, while the rest of the cells are unaffected.
A mosaic mutation may or may not be inheritable, depending on the cell populations affected. Though this is not always the case, genetic mutations can occur spontaneously in one sex cell of a parent during gametogenesis. In these cases, called germline mutations, the mutation will be present in the single-celled zygote conceived from the affected sperm or egg cell, and the condition can be inherited by the next generation.
Homologous conditions
In humans, a skin condition called piebaldism is caused by more than a dozen distinct mutations in the KIT gene. Piebaldism in humans is characterized by a white forelock, and pigmentless patches of skin on the forehead, brow, face, ventral trunk and extremities. Outside of pigmentation, piebaldism is an otherwise benign condition. In pigs, the "patch," "belted," and commercial "white" colors are caused by mutations on the KIT gene. The best-known model for KIT gene function is the mouse, in which over 90 alleles have been described. The various alleles produce everything from white toes and blazes to black-eyed white mice, panda-white to sashed and belted. Many of these alleles are lethal in the homozygous state, lethal when combined, or sublethal due to anemia. Male mice with KIT mutations are often sterile. To date, no such pleiotropic effects have been described in horses with KIT mutations.