
Phylum Chordata Order Placodont | Class Reptilia Family †Cyamodontidae Rank Genus | |
 | ||
Similar | ||
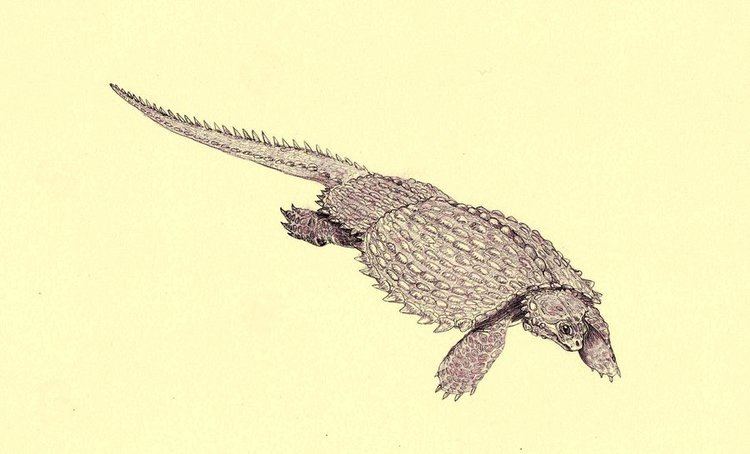
Cyamodus (pron.: SIE-ah-MO-dus) is a genus of placodonts discovered from fossil remains in Germany in the early-to-mid-19th century and was named by Christian Erich Hermann von Meyer in 1863. The fossils have been dated to the Triassic Period, from the Anisian to Ladinian stages. Considered a possible turtle ancestor due to its wide flattened torso covered in dermal ossicles, but now is considered to only be closely related. Derived from a sister to Paraplacodus, Cyamodus phylogenetically preceded Placochelys.
Contents

Meyer (1863) created the genus name Cyamodus for the "Schnabelplacodus" Cyamodus rostratus holotype skull material from Bindlach, which is today preserved and displayed in the Urwelt-Museum Oberfranken, Bayreuth.
Discovery
Cyamodus is the type genus of the monotypic family Cyamodontidae.

Thus far, five species of Cyamodus have been identified - C. rostratus, C. munsteri, C. tarnowitzensis, C. hildegardis, and C. kuhnschneyderi.

Initially considered to be an ancestral turtle due to its testudine-like head and large, bifurcated carapace. However further investigation resulted in its reclassification as a placodont, and it is closely related to other turtle-like reptiles of the Triassic period such as Henodus and Psephoderma. Similar to these other placodonts, Cyamodus made its living by hovering close to the sea floor, vacuuming up various shellfish, and grinding them between its blunt teeth.
Historically, the first Cyamodus remains were found in Upper Muschelkalk shallow marine limestones at near Bayreuth in Bavaria (Germany). They included the incomplete holotype skulls of Cyamodus muensteri and Cyamodus rostatus, which along with all other placodont remains recovered from the six quarries on the Lainecker Range in northern Bavaria were originally considered to have been derived from fish. The earliest Cyamodus skull was later restored by Muenster, with the addition of four teeth that had not been present in the original skull, and was named placodus muensteri.
Further placodont remains were found by Muenster, who collected many placodont cranial remains in the Bindlach and Lainecker Range quarries. All placodont remains from those sites were then revised as being of reptilian origin by Owen (1858). The only known complete Cyamodus skeleton, including its skull, is that of C. hildegardis, which was found outside the Germanic Basin in the northern Tethys in Switzerland. Middle Triassic sauropterygian placodonts have become increasingly important for developing new ideas to the evolutionary history of their relatives, the turtles, whereas modern analyses place placodonts not as their ancestors using morphological cladistic analyses based on the bone osteology. The study of these placodonts contributes to our understanding of the Germanic Basin and the reptile distributions.
An intriguing placodont that appears to be intermediate between Cyamodus and the placochelyids, Protenodontosaurus italicus, was described by Giovanni Pinna in 1990.
Description
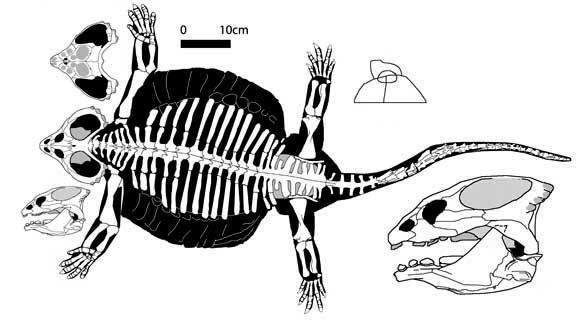
Cyamodus was a heavily armored swimmer, 1.3 meters (4.3 feet) long, that fed mainly on shellfish that it was specialized to uproot and crush with its powerful jaws. The body of Cyamodus, specifically the armor, has been described as possessing a turtle-like flatness. The shell was a two-part carapace on the upper surface of the body. The larger half covered Cyamodus from the neck to the hips and spread out flat, almost encompassing the limbs. The second, smaller plate covered the hips and the base of the tail. The shells themselves are covered in hexagonal or circular plates of armor. The skull is heart-shaped and broad.
Distinct from Paraplacodus, the skull of Cyamodus had a shorter rostrum, a smaller orbit and a larger upper temporal fenestra that was rimmed by ossifications. The teeth were flat discs, only one tooth appeared on each premaxilla and only two teeth appeared on each maxilla, with the largest teeth on the pterygoid. The quadratojugal joined the squamosal and sealed up the lateral temporal fenestra from the ancestral species (Paraplacodus).
The carapace of C. hildegardis has a series of similar-sized, enlarged lateral armor plates is rounder and less laterally expanded than was hypothesized. The separate pelvic shield, also carrying a smaller set of lateral armor plates that decrease in size with an anterior/posterior gradient covering mainly the pelvic girdle and the base of the tail. The short tail is armored by four series of armor plates that also show an anterior/posterior gradient of size reduction equivalent to the size reduction of the caudal vertebrae. Until further fossils are recovered, the internal organisation of dermal plates within the two armor shields of C. hildegardis remains little known.
Paleobiology
Placodonts were specialist eaters of shellfish and had no need to grow big and very soon were out-sized by other marine reptiles like nothosaurs. Thus, placodonts like Cyamodus developed turtle-like shells to make themselves tough for other marine reptiles to eat. Although the shells would most likely have been too cumbersome for highly adept swimming, Cyamodus would still have been more agile than other single shelled placodonts like Henodus. The shells were also covered in hexagonal plates that not only increased the level of protection but also increased their weight, a typical placodont adaptation as additional weight just beyond the level of neutral buoyancy allowed them to dive to reach shellfish. Other adaptations for protection besides the heavily armored shell include the strongly built rear of the skull and limbs that do not protrude too far. Although the reduction in limb length restricted its ability to swim, its heavy shell greatly assisted its ability to dive. Cyamodus is also expected to have had difficulty maneuvering on land and probably only ventured out of the water for egg laying and periods of rest. Discovery of two fossil Cyamodus juveniles inside the stomach area of a Lariosaurus fossil has led to speculation about its vulnerability to predation.
It has also been noted that juvenile specimens of Cyamodus have an extra tooth on the roof of their mouth, compared to adult specimens. This suggests that Cyamodus reduced the number of teeth as they grew to maturity. However, this could be due to a difference between species of Cyamodus.
Additional isolated tooth finds of C. tarnowitzensis in the Lower Muschelkalk provide evidence for the broad dispersion of an early Cyamodus species via the Carpathian Gate and later the Silesian Gate, into the central Germanic Basin where carbonate sand bar facies were widely distributed and macroalgae meadows existed. During the Middle Muschelkalk, regression and the resulting basin evaporation and facies changes caused all placodonts to disappear from the central Germanic Basin and no records are known. In the Upper Muschelkalk, C. rostatus then appeared as the next cyamodont in the Germanic Basin and by the time of the maximum marine expansion into the Germanic Basin and across the northern Tethys, this species had acquired a widespread distribution. During the Longobardian (upper Lower Keuper) full marine regression, these cyamodonts must have retreated into the macroalgae-rich environments of the northern Tethys, where they may have evolved into other species or genera of placodonts.
C. hildegardis from the Besano Formation (Middle Triassic) of the Alpine area of Switzerland and northern Italy has been reconstructed with a broad, laterally expanded main armor (carapace) and a separate smaller pelvic shield, giving it a sprawling appearance. A reexamination of the postcranial dermal armor and endoskeletal elements of the three best preserved articulated specimens of the species has led to new interpretations of the dermal armor and underlying postcranial bones, as well as a new life reconstruction.
Geological and Environmental Information
The oldest record of Cyamodus is a skull of C. tarnowitzensis from Pelsonian shallow marine deposits. During the middle Illyrian, placodonts disappeared from the Germanic Basin. With renewed upper Illyrian transgression, C. rostratus was found in terebratulid shell-rich shallow marine deposits. Abundant remains of Cyamodus muensteri have been reported from the upper Illyrian/middle Fassanian. Skeletal remains of this species are from the Grenzbitumenzone of the Monte San Giorgio lagoons. The most recent species, C. kuhnschnyderi, has been found in the upper Fassanian/lower Longobardian of the southern Germanic Basin or Burgundian Gate when marine facies in the Germanic Basin had nearly disappeared. These successive species provide evidence of monophylogenetic development with a trend towards anterior upper and lower jaw teeth reduction, along with a shortening of the rostrum (over interval of five million years, 243-238 Ma), suggesting an evolutionary adaptation to specialized feeding.
The sediments of the Germanic Basin which contain Cyamodus and other placodont remains are generally similar in both the Lower and Upper Muschelkalk, comprising terebratulid and shell-rich shallow subtidal partly oolithic carbonates.
Cyamodus remains were absent from the Germanic Basin during the Middle Muschelkalk, when tectonically controlled uplift resulted in shallowing of the basin, evaporation and a basin-wide disappearance of the macroalgae meadows.
Evolution
The observed pattern of tooth reductions provides evidence supporting a newer interpretation that placodonts grazed on macroalgae, indicating that Cyamodus changed from an omnivore to an algae-grazer. The late/Upper Muschelkalk cyamodont C. kuhnschnyderi represents the most highly evolved Cyamodus species in the Germanic Basin, with the shortest rostrum, the greatest tooth reduction (having just one premaxillary, two maxillary and two palatine teeth) and the greatest tooth enlargement in both the upper and the lower jaws. The lower jaws still have three teeth, but now they also consist of the broadest anterior teeth of all the cyamodonts. Overall, this species has a dentition type similar to that of Placodus, with a food-squeezing function for both the lower and the upper jaws.
Cyamodus did not have any dorsal spines, although it did have a wing-like elongated flattened lateral spine that served to brace the overlying subdermal carapace. Presumably, the dorsal spines disappeared to provide a closer and better braced association between the vertebrae and the carapace.