
Symbol MCD1 Entrez 851561 UniProt Q12158 | Alt. symbols PDS3, RHC21, SCC1 PDB 1W1W | |
 | ||
Cohesin and condensin
Cohesin is a protein complex that regulates the separation of sister chromatids during cell division, either mitosis or meiosis. Cohesins hold sister chromatids together after DNA replication until anaphase when removal of cohesin leads to separation of sister chromatids.
Contents
- Cohesin and condensin
- Kim nasmyth how does cohesin hold dnas together
- Structure
- Function
- Dissociation of sister chromatid cohesion
- Mechanism of action
- Localization of cohesin rings
- Evolution
- Clinical significance
- References
Kim nasmyth how does cohesin hold dnas together
Structure
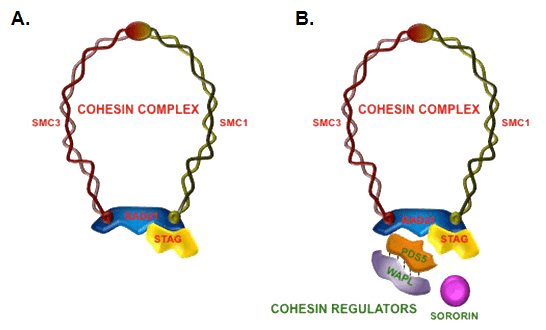
Cohesin is a multi-subunit protein complex, made up of four core subunits: two SMC proteins (SMC1 and 3), an alpha-kleisin (orthologues of yeast Scc1), and an orthologue of the yeast Scc3 protein (e.g. STAG1-3 in humans and SA1-3 in mice).
Smc1 and Smc3 are members of the Structural Maintenance of Chromosomes (SMC) family. SMC proteins have two main structural characteristics: an ATP-binding cassette-like 'head' domain with ATPase activity (formed by the interaction of the N- and C- terminals) and a hinge domain that allows dimerization of SMCs. The head and the hinge domains are connected to each other via long anti-parallel coiled coils. The dimer is present in a V-shaped form, connected by the hinges. Upon ATP binding, the two head domains in the dimer bind to each other, forming a ring structure. ATP hydrolysis can therefore trigger opening and closing of the ring.
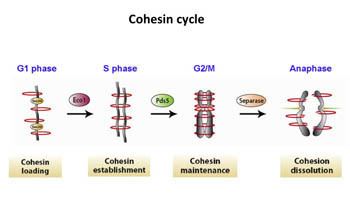
Scc1 and Scc3 bind the ATPase domains of Smc1 and Smc3 stabilizing the ring structure. The amino and carboxy terminus of Scc1 bind Smc1 and Smc3. Once Scc1 binds the SMC proteins, Scc3 can also associate by binding with the C-terminal region of Scc1. When Scc1 binds on both Smc1 and Smc3, the cohesin complex forms a closed ring structure. When it binds to only one of the SMC proteins, the complex forms an open ring. Early studies suggested various ways in which cohesin may entrap DNA, including as a monomer that holds both homologues together, and a "hand-cuff" model where two intertwining cohesin complexes each hold one sister chromatid. While some studies support the idea of a hand-cuff model, the model is inconsistent with a number of experimental observations, and is generally considered to entrap chromatin as a monomer.
The topology and structure of these subunits has been best characterized in budding yeast (Haering et al. 2002, 2004), but the sequence conservation of these proteins and biochemical and electron microscopic observations imply that cohesin complexes in other species are very similar in their structure, [1].
Function
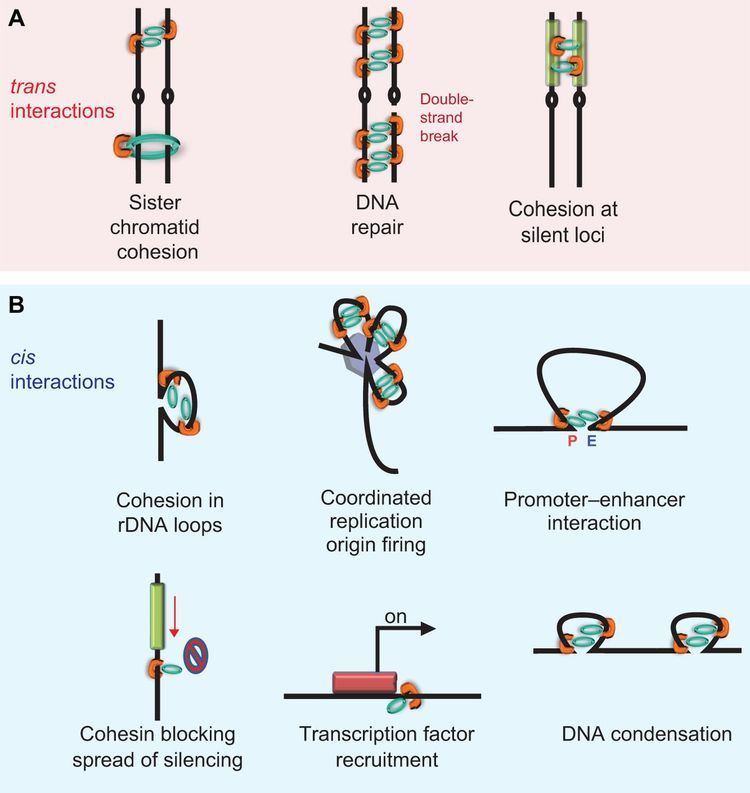
The cohesin ring has many functions:
1. It is used to keep the sister chromatids connected with each other during metaphase ensuring that during mitosis (and meiosis), each sister chromatid segregates to opposite poles. Without cohesin, the cell would be unable to control sister chromatid segregation since there would be no way of ensuring whether the spindle fiber attached on each sister chromatid is from a different pole.
2. It facilitates spindle attachment onto chromosomes.
3. It facilitates DNA repair by recombination.
4. Recently many novel functions of cohesin have been discovered in many different cellular processes. Cohesin has been shown to be responsible for transcription regulation, DNA double strand break repair, chromosome condensation, pairing of homologous chromosomes during meiosis I, mono-orientation of sister kinetochores during meiosis I, non-homologous centromere coupling, chromosome architecture and rearrangement, DNA replication etc.
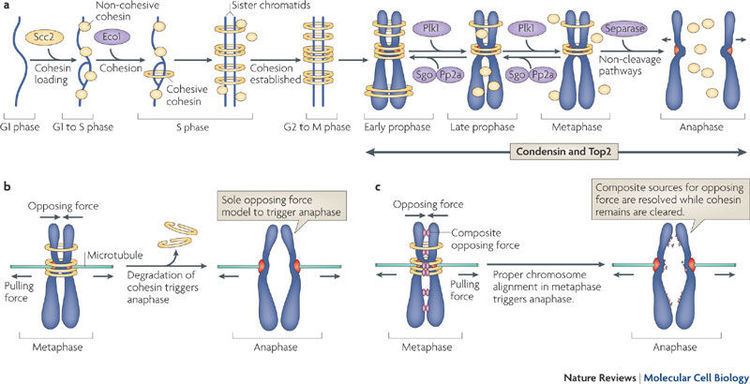
Dissociation of sister chromatid cohesion
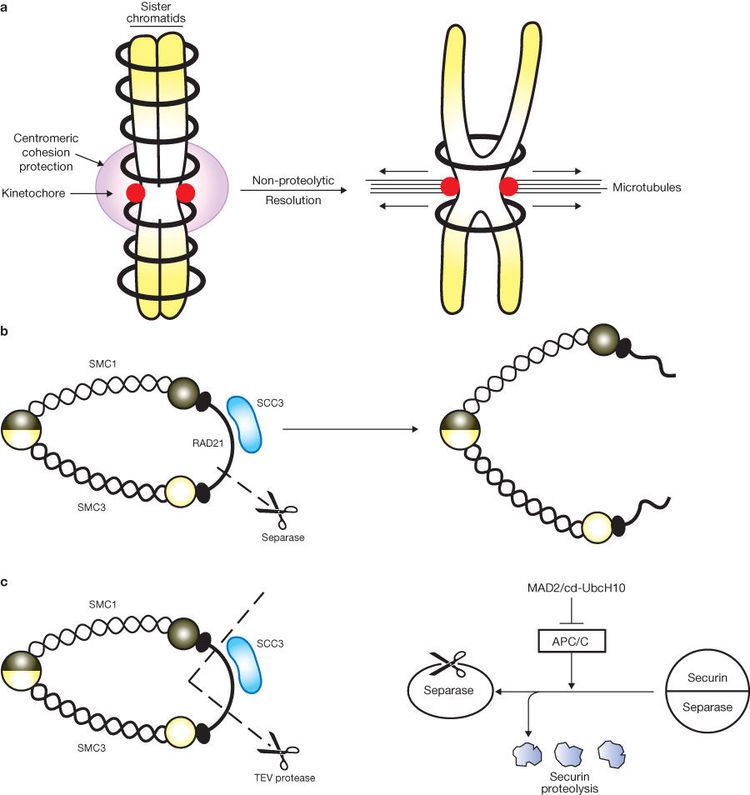
The anaphase promoting complex associated to Cdc20 (APC/C-cdc20) cleaves the securin (anaphase inhibitor), which holds the sister chromatids together. Securin is cleaved at anaphase, following APC/C-cdc20 mediated degradation, and it renders separase (a protease, inhibited by the association with securin) to cleave the kleisin subunit. An alpha-kleisin is associated with the cohesin complex, linking both SMC 3 and SMC 1 together, with the exact kleisin varying between mitosis and meiosis (Scc1 and Rec8 respectively), and its cleavage ultimately leads to the removal of cohesin from chromosomes.
Dissociation of sister chromatids cohesion defines anaphase onset, which establishes two sets of identical chromosomes at each pole of the cell (telophase). Then the two daughter cells separate, and a new round of the cell cycle freshly starts in each one, at the stage of G0. When cells are ready to divide, because cell size is big enough or because they receive the appropriate stimulus, they activate the mechanism to enter into the G1 stage of cell cycle, and they duplicate most organelles during S (synthesis) phase, including their centrosome. Therefore, when the cell division process will end, each daughter cell will receive a complete set of organelles. At the same time, during S phase all cells must duplicate their DNA very precisely, a process termed DNA replication. Once DNA replication has finished, in eukaryotes the DNA molecule is compacted and condensed, to form the mitotic chromosomes, each one constituted by two sister chromatids, which stay held together by the establishment of cohesion between them; each chromatid is a complete DNA molecule, attached via microtubules to one of the two centrosomes of the dividing cell, located at opposed poles of the cell. To avoid premature sister chromatid separation, the APC/C is maintained in an inactive state bound to different molecules, which are part of a complex mechanism termed the spindle assembly checkpoint.
Mechanism of action
It is not clear how the cohesion ring links sister chromatids together. There are two possible scenarios:
- Cohesin subunits bind to each sister chromatid and form a bridge between the two.
- Since cohesin has a ring structure, it is able to encircle both sister chromatids.
Current evidence suggests that the second scenario is the most likely. Proteins that are essential for sister chromatid cohesion, such as Smc3 and Scc1, do not regulate the formation of covalent bonds between cohesin and DNA, indicating that DNA interaction is not sufficient for cohesion. In addition, disturbing the ring structure of cohesin through cleavage of Smc3 or Scc1 triggers premature sister chromatid segregation in vivo. This shows that the ring structure is important for cohesin’s function.
Even though the ring hypothesis appears to be valid, there are still questions about the number of rings required to hold sister chromatids together. One possibility is that one ring surrounds the two chromatids. Another possibility involves the creation of a dimer where each ring surrounds one sister chromatid. The two rings are connected to each other through formation of a bridge that holds the two sister chromatids together.
The cohesion complex is established during the initial stages of S-phase. The complexes associate with chromosomes before DNA replication occurs. Once cells start replicating their DNA, cohesin rings close and link the sister chromatids together. Cohesin complexes must be present during S-phase in order for cohesion to take place. It is unclear, however, how cohesin is loaded on the chromosomes during G1. There are two proposed hypotheses so far:
- The ATPase domain of the SMC proteins interacts with DNA and this interaction initially mediates the loading of cohesin complexes on chromosomes.
- Several proteins aid in the loading process. For example, Scc2 and Scc4 are both required for cohesin to load in budding yeast.
Localization of cohesin rings
Cohesin binding along the chromosomal DNA is considered to be dynamic and its location changes based on gene transcription, specific DNA sequence and presence of chromosome-associated proteins. There are three possible scenarios:
- Cohesin location is influenced by the orientation of neighboring genes and it is most frequently located in areas of convergent transcription. Gene orientation depends on the direction of transcription and can be of three types: head-to-head, head-to-tail and tail-to-tail. The tail-to-tail configuration results in the convergence of transcription machinery. One hypothesis states that the RNA polymerase “pushes” cohesin along the DNA, causing them to move towards the direction of the RNA polymerases. Changing the transcription pattern of genes changes the location of cohesin indicating that localization of cohesin may depend on transcription.
- A few cohesin rings are found in chromosome arms that have AT-rich DNA sequences indicating that DNA sequence may be an independent factor of cohesin binding.
- Cohesin rings, especially in budding yeast, are also located in the region surrounding the centromere. Two hypotheses may explain this: the presence of repetitive heterochromatic DNA in centromeres and the presence of chromosome-associated proteins. For example, Schizosaccharomyces pombe have multiple copies of specific heterochromatic DNA whose involvement in cohesion binding has been proven. Budding yeast lacks repetitive sequences and, therefore, requires a different mechanism for cohesion binding. Evidence suggests that binding of cohesin to the budding yeast centromere region depends on chromosome-associated proteins of the kinetochore that mediate cohesion association to pericentric regions (the kinetochore is an enhancer of pericentric cohesin binding).
Evolution
Cohesin structure and function has been conserved in evolution. The SMC proteins are found in prokaryotes and have been conserved through evolution. The coiled coils of SMC1 and SMC3 are conserved with an amino acid divergence of less than 0.5%.
Clinical significance
The term "cohesinopathy" has been used to describe conditions affecting the cohesin complex.
These conditions include: