
 | ||
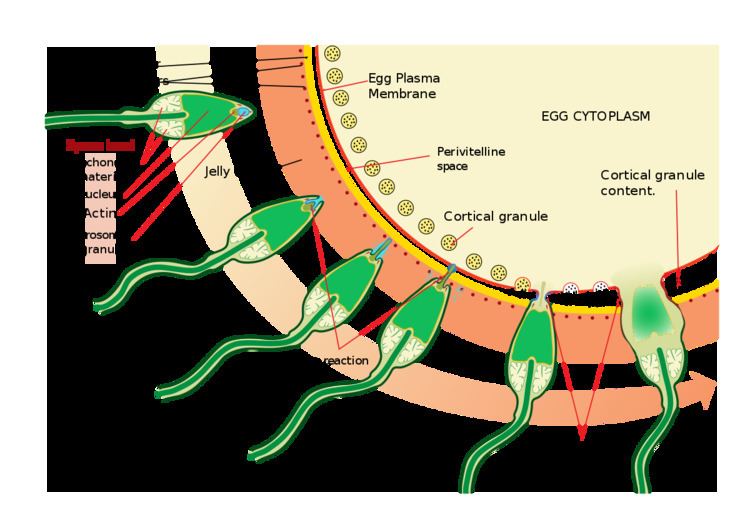
During fertilization, a sperm must first fuse with the plasma membrane and then penetrate the female egg in order to fertilize it. Fusing to the egg usually causes little problem, whereas penetrating through the egg's hard shell or extracellular matrix can present more of a problem to the sperm. Therefore, sperm cells go through a process known as the acrosome reaction which is the reaction that occurs in the acrosome of the sperm as it approaches the egg. The acrosome is a cap-like structure over the anterior half of the sperm's head.
Contents
As the sperm approaches the zona pellucida of the egg, which is necessary for initiating the acrosome reaction, the membrane surrounding the acrosome fuses with the plasma membrane of the sperm's head, exposing the contents of the acrosome. The contents include surface antigens necessary for binding to the egg's cell membrane, and numerous enzymes which are responsible for breaking through the egg's tough coating and allowing fertilization to occur.
Variations among species
There are considerable species variations in the morphology and consequences of the acrosome reaction. In several species the trigger for the acrosome reaction has been identified in a layer that surrounds the egg.
Echinoderms
In some lower animal species a protuberance (the acrosomal process) forms at the apex of the sperm head, supported by a core of actin microfilaments. The membrane at the tip of the acrosomal process fuses with the egg's plasma membrane.
In some echinoderms, including starfish and sea urchins, a major portion of the exposed acrosomal content contains a protein that temporarily holds the sperm on the egg's surface.
Mammals
In mammals the acrosome reaction releases hyaluronidase and acrosin; their role in fertilization is not yet clear. The acrosomal reaction does not begin until the sperm comes into contact with the oocyte's zona pellucida. Upon coming into contact with the zona pellucida, the acrosomal enzymes begin to dissolve and the actin filament comes into contact with the zona pellucida. Once the two meet, a calcium influx occurs, causing a signaling cascade. The cortical granules inside the oocyte then fuse to the outer membrane and a transient fast block reaction occurs.
It also alters a patch of pre-existing sperm plasma membrane so that it can fuse with the egg plasma membrane.
A sperm penetration assay includes an acrosome reaction test that assesses how well a sperm is able to perform during the fertilization process. Sperm that are unable to properly go through the acrosome reaction will not be able to fertilize an egg. However, this problem only occurs in about 5% of men that have the test done. This test is rather expensive and provides limited information on a man's fertility.
In other cases, such as in the wood mouse Apodemus sylvaticus, premature acrosome reactions have been found to cause increased motility in aggregates of spermatozoa promoting fertilization
The process
The acrosomal reaction normally takes place in the ampulla of the fallopian tube (site of fertilization) when the sperm penetrates the secondary oocyte. A few events precede the actual acrosome reaction. The sperm cell acquires a "hyperactive motility pattern" by which its flagellum produces vigorous whip-like movements that propel the sperm through the cervical canal and uterine cavity, until it reaches the isthmus of the fallopian tube. The sperm approaches the ovum in the ampulla of the fallopian tube with the help of various mechanisms, including chemotaxis. Glycoproteins on the outer surface of the sperm then bind with glycoproteins on the zona pellucida of the ovum.
The first stage is the penetration of corona radiata, by releasing hyaluronidase from the acrosome to digest cumulus cells surrounding the oocyte and exposing acrosin attached to the inner membrane of the sperm. The cumulus cells are embedded in a gel-like substance made primarily of hyaluronic acid, and developed in the ovary with the egg and support it as it grows. After reaching the zona pellucida the actual acrosome reaction begins.
Acrosin digests the zona pellucida and membrane of the oocyte. Part of the sperm's cell membrane then fuses with the egg cell's membrane, and the contents of the head sink into the egg. In the mouse it has been demonstrated that ZP3, one of the proteins that make up the zona pellucida, binds to a partner molecule (to the β1,4-galactosyl transferase receptors) on the sperm. This lock-and-key type mechanism is species-specific and prevents the sperm and egg of different species from fusing. The zona pellucida also releases Ca granules to prevent additional sperm from binding. There is some evidence that this binding is what triggers the acrosome to release the enzymes that allow the sperm to fuse with the egg. It is likely that a similar mechanism occurs in other mammals, but the diversity of zona proteins across species means that the relevant protein and receptor may differ.
Upon penetration, if all is occurring normally, the process of egg-activation occurs and the oocyte is said to have become activated. This is thought to be induced by a specific protein phospholipase c zeta. It undergoes its secondary meiotic division, and the two haploid nuclei (paternal and maternal) fuse to form a zygote. In order to prevent polyspermy and minimise the possibility of producing a triploid zygote, several changes to the egg's cell membranes renders them impenetrable shortly after the first sperm enters the egg.
The aforementioned process describes the physiologically relevant events. One should however bear in mind that a certain percentage of sperm cells will undergo a spontaneous acrosome reaction without the presence of the ovum. Those cells are not able to fertilise the egg, even if they do reach it later. Other cells will spontaneously shed their acrosome during the process of apoptosis/necrosis.
In in vitro fertilization
When using intracytoplasmic sperm injection (ICSI) for IVF, the implantation rate is higher in oocytes injected with spermatozoa that have undergone acrosome reaction (~40%) vs. those injected with nonreacted spermatozoa (~10%). The implantation rate is ~25% in when injected with both reacted and nonreacted spermatozoa. The delivery rate per cycle follows the same trend.
The acrosome reaction can be stimulated in vitro by substances a sperm cell may encounter naturally such as progesterone or follicular fluid, as well as the more commonly used calcium ionophore A23187.
Assessment
Birefringence microscopy, flow cytometry or fluorescence microscopy can be used for assessing the shedding of the acrosome or "acrosome reaction" of a sperm sample. Flow cytometry and fluorescence microscopy are usually done after staining with a fluoresceinated lectin such as FITC-PNA, FITC-PSA, FITC-ConA, or fluoresceinated antibody such as FITC-CD46. The antibodies/lectins have a high specificity for different parts of the acrosomal region, and will only bind to a specific site (acrosomal content/ inner/outer membrane). If bound to a fluorescent molecule, regions where these probes have bound can be visualised. Sperm cells with artificially induced acrosome reactions may serve as positive controls.
For fluorescence microscopy a smear of washed sperm cells are made, airdried, permealized and then stained. Such a slide is then viewed under light of a wavelength that will cause the probe to fluoresce if it is bound to the acrosomal region. At least 200 cells are viewed in an arbitrary fashion and classified as either acrosome intact (fluorescing bright green) or acrosome reacted (no probe present, or only on the equatorial region). It is then expressed as a percentage of the counted cells.
For assessment with flow cytometry the washed cells are incubated with the chosen probe, (possibly washed again) and then sampled in a flow cytometer. After gating the cell population according to forward- and side-scatter the resulting data can be analysed (E.g. mean fluorescences compared). With this technique a probe for viability, like propidium iodide (PI) could also be included in order to exclude dead cells from the acrosome assessment, since many sperm cells will spontaneously lose their acrosome when they die.