
Latin organum vomeronasale TA A06.1.02.008 | Lymph Node Dorlands/Elsevier 12596317 FMA 77280 | |
 | ||
The vomeronasal organ (VNO), or the Jacobson's organ, is an auxiliary olfactory sense organ that is found in many animals. It lies close to the vomer and nasal bones. It was discovered by Frederik Ruysch prior to 1732 and later by Ludwig Jacobson in 1813. This organ is the sense organ involved in the flehmen response in mammals.
Contents
- Structure
- Function
- Sensory epithelium and receptors
- Sensory neurons
- Animals
- Behavioral studies
- Humans
- References
The VNO is the first stage of the accessory olfactory system and contains sensory neurons that detect chemical stimuli. The axons from these neurons project to the accessory olfactory bulb, which targets the amygdala and bed nucleus of the stria terminalis, which in turn project to the hypothalamus.
The vomeronasal organ is mainly used to detect pheromones, chemical messengers that carry information between individuals of the same species. As with other olfactory systems, chemical messages are detected by their binding to G protein-coupled receptors. The neurons in the VNO express receptors from three families, called V1R, V2R, and FPR. The receptors are distinct from each other and from the large family of receptors in the main olfactory system. Stimuli reach the VNO in liquid phase via a pumping mechanism; the primary cues for the VNO are therefore non-volatile and require direct physical contact.
Its presence in many animals has been widely studied and the importance of the vomeronasal system to the role of reproduction and social behavior (through its influence on the anterior hypothalamus) has been shown in many studies. Its presence and functionality in humans was controversial, though most studies agree the organ regresses during fetal development. Many genes essential for VNO function in animals (such as TRPC2) are non-functional in humans. Chemical communication does appear to occur among humans, but this does not necessarily imply that the human vomeronasal organ is functional.
Structure
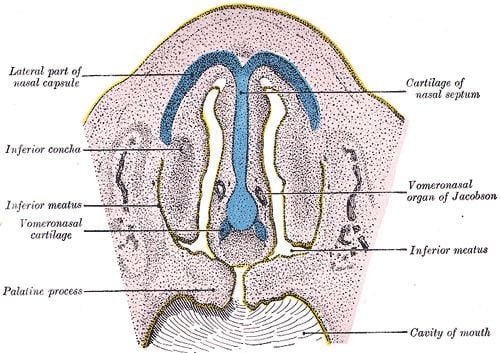
The VNO is found at the base of the nasal cavity. It is split into two, being divided by the nasal septum, with both sides possessing an elongated C-shaped, or crescent, lumen. It is encompassed inside a bony or cartilaginous capsule which opens into the base of the nasal cavity. The vomeronasal receptor neurons possess axons which travel from the VNO to the accessory olfactory bulb (AOB) or, as it's also known, the vomeronasal bulb. These sensory receptors are located on the medial concave surface of the crescent lumen. The lateral, convex surface of the lumen is covered with non-sensory ciliated cells, where the basal cells are also found. At the dorsal and ventral aspect of the lumen are vomeronasal glands, which fill the vomeronasal lumen with fluid. Sitting next to the lumen are blood vessels that dilate or constrict, forming a vascular pump that deliver stimuli to the lumen. A thin duct, which opens onto the floor of the nasal cavity inside the nostril, is the only way of access for stimulus chemicals.
During embryological development, the vomeronasal sensory neurons form from the nasal (olfactory) placode, at the anterior edge of the neural plate (cranial nerve zero).
Function
In mammals, the sensory neurons of the vomeronasal organ detect non-volatile chemical cues, which requires direct physical contact with the source of odor. Notably, some scents act as chemical-communication signals (pheromones) from other individuals of the same species. Unlike the main olfactory bulb that sends neuronal signals to the olfactory cortex, the VNO sends neuronal signals to the accessory olfactory bulb and then to the amygdala, BNST, and ultimately hypothalamus. Since the hypothalamus is a major neuroendocrine center (affecting aspects of reproductive physiology and behavior as well as other functions such as body temperature), this may explain how scents influence aggressive and mating behavior. For example, in many vertebrates, nerve signals from the brain pass sensory information to the hypothalamus about seasonal changes and the availability of a mate. In turn, the hypothalamus regulates the release of reproductive hormones required for breeding. It should be noted that some pheromones are detected by the main olfactory system.
Sensory epithelium and receptors
The VNO is a tubular crescent shape and split into two pairs, separated by the nasal septum. The medial, concave area of the lumen is lined with a pseudo stratified epithelium that has three main cell types: receptor cells, supporting cells, and basal cells. The supporting cells are located superficially on the membrane while the basal cells are found on the basement membrane near the non-sensory epithelium. The receptor neurons possess apical microvilli, to which are localized the sensory receptors, G-protein-coupled receptors which are often referred to as pheromone receptors since vomeronasal receptors have been tied to detecting pheromones.
There have been three different G-protein-coupled receptors identified in the VNO, each found in distinct regions: the V1Rs, V2Rs, and FPRs. V1Rs, V2Rs and FPRs are seven transmembrane receptors which are not closely related to odorant receptors expressed in the main olfactory neuroepithelium.
The vomeronasal organ’s sensory neurons act on a different signaling pathway than that of the main olfactory system’s sensory neurons. Activation of the receptors stimulates phospholipase C, which in turn opens the ion channel TRPC2. Upon stimulation activated by pheromones, IP3 production has been shown to increase in VNO membranes in many animals, while adenylyl cyclase and cyclic adenosine monophosphate (cAMP), the major signaling transduction molecules of the main olfactory system, remain unaltered. This trend has been shown in many animals, such as the hamster, the pig, the rat, and the garter snake upon introduction of vaginal or seminal secretions into the environment.
V1Rs and V2Rs are suggested to be activated by distinct ligands or pheromones. The evidence that Gi and Go proteins are activated upon stimulation via different pheromones supports this.
Many vomeronasal neurons are activated by chemicals in urine. Some of the active compounds are sulfated steroids. Detecting the types and amounts of different sulfated steroids conveys information about the urine donor's physiological state, and may therefore serve as an honest signal.
Recent studies proved a new family of formyl peptide receptor like proteins in VNO membranes of mice, which points to a close phylogenetic relation of signaling mechanisms used in olfaction and chemotaxis.
Sensory neurons
Vomeronasal sensory neurons are extremely sensitive and fire action potentials at currents as low as 1 pA. Many patch-clamp recordings have confirmed the sensitivity of the vomeronasal neurons. This sensitivity is tied to the fact that the resting potential of the vomeronasal neurons is relatively close to that of the firing threshold of these neurons. Vomeronasal sensory neurons also show remarkably slow adaptation and the firing rate increases with increasing current up to 10 pA. The main olfactory sensory neurons fire single burst action potentials and show a much quicker adaptation rate. Activating neurons that have V1 receptors, V1Rs, cause field potentials that have weak, fluctuating responses that are seen the anterior of the accessory olfactory bulb, AOB. Activation of neurons that contain V2 receptors, V2Rs, however, promote distinct oscillations in the posterior of the AOB.
Animals
The functional vomeronasal system is found in many animals, including all snakes and lizards, plus many mammals, such as mice, rats, elephants, cattle, dogs, cats, goats and pigs.
In some other mammals the entire organ contracts or pumps in order to draw in the scents.
Some mammals, particularly felids (cats) and ungulates (which includes horses, cattle, and pigs among other species), use a distinctive facial movement called the Flehmen response to direct inhaled compounds to this organ. The animal will lift its head after finding the odorant, wrinkle its nose while lifting its lips, and cease to breathe momentarily. Flehmen behavior is associated with “anatomical specialization”, and animals that present flehmen behavior have incisive papilla and ducts, which connect the oral cavity to the VNO, that are found behind their teeth. However, horses are the exception: they exhibit Flehmen response but do not have an incisive duct communication between the nasal and the oral cavity because they do not breathe through their mouths, instead, the VNOs connect to the nasal passages by the nasopalatine duct.
Behavioral studies
Kudjakova et al. performed exploratory behavioral studies of non purebred rats by extirpating the VNO. The study showed that the exploratory behavior of the rats with extirpated VNO’s were significantly different from both control groups of rats. These results suggest that removal of the VNO removed the experimental rats from important social information. This is seen in the reduced exploratory activity in the experimental animal and the lower number of species-specific reactions.
Another study conducted by Beauchamp et al. investigated the role of the VNO in male guinea pigs social behavior. Half of the guinea pigs systems were removed, while the other half were put under fake surgeries with their vomeronasal systems left intact. The findings suggested that the VNO in the male domestic guinea pig is necessary for the maintenance of normal responsiveness to sex odors. However, “in its absence, other sensory systems are capable of maintaining normal sexual behavior under conditions of laboratory testing.”
Studies of species with a vomeronasal organ (VNO) have demonstrated its importance in the detection of and response to pheromones. Studies have shown that the VNO responds primarily to socially relevant pheromonal cues. Removal of the VNO impairs and, in many cases, causes the loss of sexual behavior in male rats. Further, lesioning the VNO results in an increase in the latency to the first intromission and ejaculation. Lesions also cause a decrease in the number of ejaculations and their efficiency. Sexually inexperienced rats that have lesions of the VNO have more impairments in sexual behavior than controls. These data contrast with similar studies in hamsters, that found that removal of the VNO in sexually experienced males had little to no effect behavior. Within rats, the VNO is sexually dimorphic, being larger in males than females. However, there is no discernible difference in size between control males and androgenized females or control females and castrated males, provided the androgenization and castration are prepubescent. These data suggests that testosterone is responsible for development of the VNO.
These behavioral studies show the importance of the vomeronasal system in animals’ social networks and everyday activities. The importance of the vomeronasal system to the role of reproduction and social networking has been shown in many studies. Neuromodulatory systems can target the vomeronasal system to mediate sexual behaviors signaled by the accessory olfactory bulb
Humans
Many studies have been performed to determine whether there is an actual presence of a VNO in adult human beings. Trotier et al. estimated that around 92% of their subjects that had no septal surgery had at least one intact VNO. Kjaer and Fisher Hansen, on the other hand, stated that VNO structure disappeared during fetal development as it does for some primates. However, Smith and Bhatnagar (2000) asserted that Kjaer and Fisher Hansen simply missed the structure in older fetuses. Won (2000) found evidence of a VNO in 13 of his 22 cadavers (59.1%) and in 22 of his 78 living patients (28.2%). In a study published in 2016 Stoyanov et al. found the organ to be present in 26.83% of the Bulgarian population, using retrospective analysis of nearly a thousand outpatient nasal endoscopies.
Given these findings, some scientists have argued that there is a VNO in adult human beings. However, most investigators have sought to identify the opening of the vomeronasal organ in humans, rather than identify the tubular epithelial structure itself. Thus it has been argued that such studies, employing macroscopic observational methods, have sometimes missed or even misidentified the vomeronasal organ.[21]
Among studies that use microanatomical methods, there is no reported evidence that human beings have active sensory neurons like those in working vomeronasal systems of other animals.[21] Furthermore, there is no evidence to date that suggests there are nerve and axon connections between any existing sensory receptor cells that may be in the adult human VNO and the brain. Likewise, there is no evidence for any accessory olfactory bulb in adult human beings, and the key genes involved in VNO function in other mammals have pseudogeneized in human beings. Therefore, while the presence of a structure in adult human beings is debated, a review of the scientific literature by Tristram Wyatt concluded, "most in the field... are sceptical about the likelihood of a functional VNO in adult human beings on current evidence."