
 | ||
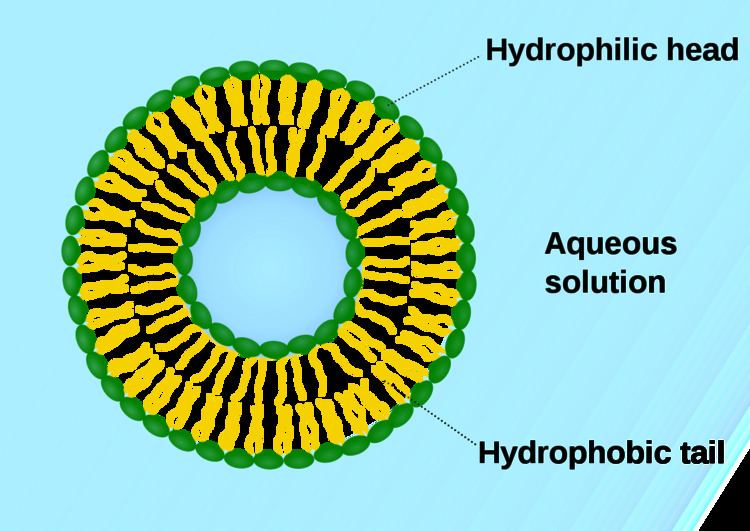
In cell biology, a vesicle is a small structure within a cell, consisting of fluid enclosed by a lipid bilayer. Vesicles form naturally during the processes of secretion (exocytosis), uptake (endocytosis) and transport of materials within the cytoplasm. Alternatively, they may be prepared artificially, in which case they are called liposomes. (Not lysosomes) If there is only one phospholipid bilayer, they are called unilamellar liposome vesicles; otherwise they are called multilamellar. The membrane enclosing the vesicle is also a lamellar phase, similar to that of the plasma membrane and vesicles can fuse with the plasma membrane to release their contents outside the cell. Vesicles can also fuse with other organelles within the cell.
Contents
- Vacuoles
- Lysosomes
- Transport vesicles
- Secretory vesicles
- Types
- Extracellular vesicles
- Other types
- Vesicle formation and transport
- Vesicle coat and cargo molecules
- Vesicle docking
- Vesicle fusion
- Vesicles in receptor downregulation
- Isolated vesicles
- Artificial vesicles
- References
Vesicles perform a variety of functions. Because it is separated from the cytosol, the inside of the vesicle can be made to be different from the cytosolic environment. For this reason, vesicles are a basic tool used by the cell for organizing cellular substances. Vesicles are involved in metabolism, transport, buoyancy control, and temporary storage of food and enzymes . They can also act as chemical reaction chambers.
The 2013 Nobel Prize in Physiology or Medicine was shared by James Rothman, Randy Schekman and Thomas Südhof for their roles (building upon earlier research, some of it by their mentors) on the makeup and function of cell vesicles, especially in yeasts and in humans, including information on each vesicle's parts and how they are assembled. Vesicle dysfunction is thought to contribute to Alzheimer's disease, diabetes, some hard-to-treat cases of epilepsy, some cancers and immunological disorders and certain neurovascular conditions.
Vacuoles
Vacuoles are vesicles which contain mostly water.
Lysosomes
Transport vesicles
Secretory vesicles
Secretory vesicles contain materials that are to be excreted from the cell. Cells have many reasons to excrete materials. One reason is to dispose of wastes. Another reason is tied to the function of the cell. Within a larger organism, some cells are specialized to produce certain chemicals. These chemicals are stored in secretory vesicles and released when needed.
Types
Extracellular vesicles
Extracellular vesicles (EVs) are produced by all domains of life including complex eukaryotes, both Gram-negative and Gram-positive bacteria, mycobacteria and fungi.
Types
These are often separated by density by differential centrifugation.
Ectosomes were named in 2008, but in 2012 are not considered a separate type.
In humans, endogenous extracellular vesicles likely play a role in coagulation, intercellular signaling and waste management.
In Gram-negative bacteria, EVs are produced by the pinching off of the outer membrane; however, how EVs escape the thick cell walls of Gram-positive bacteria, mycobacteria and fungi is still unknown. These EVs contain varied cargo, including nucleic acids, toxins, lipoproteins and enzymes and have important roles in microbial physiology and pathogenesis. In host-pathogen interactions, gram negative bacteria produce vesicles which play roles in establishing a colonization niche, carrying and transmitting virulence factors into host cells and modulating host defense and response.
Ocean cyanobacteria have been found to continuously release vesicles containing proteins, DNA and RNA into the open ocean. Vesicles carrying DNA from diverse bacteria are abundant in coastal and open-ocean seawater samples.
Other types
Gas vesicles are used by Archaea, bacteria and planktonic microorganisms, possibly to control vertical migration by regulating the gas content and thereby buoyancy, or possibly to position the cell for maximum solar light harvesting. These vesicles are typically lemon-shaped or cylindrical tubes made out of protein; their diameter determines the strength of the vesicle with larger ones being weaker. The diameter of the vesicle also affects it's volume and how efficiently it can provide buoyancy. In cyanobacteria natural selection has worked to create vesicles that are at the maximum diameter possible while still being structurally stable. The protein skin is permeable to gasses but not water, keeping the vesicles from flooding.
Matrix vesicles are located within the extracellular space, or matrix. Using electron microscopy they were discovered independently in 1967 by H. Clarke Anderson and Ermanno Bonucci. These cell-derived vesicles are specialized to initiate biomineralisation of the matrix in a variety of tissues, including bone, cartilage and dentin. During normal calcification, a major influx of calcium and phosphate ions into the cells accompanies cellular apoptosis (genetically determined self-destruction) and matrix vesicle formation. Calcium-loading also leads to formation of phosphatidylserine:calcium:phosphate complexes in the plasma membrane mediated in part by a protein called annexins. Matrix vesicles bud from the plasma membrane at sites of interaction with the extracellular matrix. Thus, matrix vesicles convey to the extracellular matrix calcium, phosphate, lipids and the annexins which act to nucleate mineral formation. These processes are precisely coordinated to bring about, at the proper place and time, mineralization of the tissue's matrix unless the Golgi are non-existent.
Multivesicular body, or MVB, is a membrane-bound vesicle containing a number of smaller vesicles.
Vesicle formation and transport
Some vesicles are made when part of the membrane pinches off the endoplasmic reticulum or the Golgi complex. Others are made when an object outside of the cell is surrounded by the cell membrane.
Vesicle coat and cargo molecules
The vesicle "coat" is a collection of proteins that serve to shape the curvature of a donor membrane, forming the rounded vesicle shape. Coat proteins can also function to bind to various transmembrane receptor proteins, called cargo receptors. These receptors help select what material is endocytosed in receptor-mediated endocytosis or intracellular transport.
There are three types of vesicle coats: clathrin, COPI and COPII. The various types of coat proteins help with sorting of vesicles to their final destination. Clathrin coats are found on vesicles trafficking between the Golgi and plasma membrane, the Golgi and endosomes and the plasma membrane and endosomes. COPI coated vesicles are responsible for retrograde transport from the Golgi to the ER, while COPII coated vesicles are responsible for anterograde transport from the ER to the Golgi.
The clathrin coat is thought to assemble in response to regulatory G protein. A protein coat assembles and disassembles due to an ADP ribosylation factor (ARF) protein.
Vesicle docking
Surface proteins called SNAREs identify the vesicle's cargo and complementary SNAREs on the target membrane act to cause fusion of the vesicle and target membrane. Such v-SNARES are hypothesised to exist on the vesicle membrane, while the complementary ones on the target membrane are known as t-SNAREs.
Often SNAREs associated with vesicles or target membranes are instead classified as Qa, Qb, Qc, or R SNAREs owing to further variation than simply v- or t-SNAREs. An array of different SNARE complexes can be seen in different tissues and subcellular compartments, with 36 isoforms currently identified in humans.
Regulatory Rab proteins are thought to inspect the joining of the SNAREs. Rab protein is a regulatory GTP-binding protein and controls the binding of these complementary SNAREs for a long enough time for the Rab protein to hydrolyse its bound GTP and lock the vesicle onto the membrane.
Vesicle fusion
Vesicle fusion can occur in one of two ways: full fusion or kiss-and-run fusion. Fusion requires the two membranes to be brought within 1.5 nm of each other. For this to occur water must be displaced from the surface of the vesicle membrane. This is energetically unfavorable and evidence suggests that the process requires ATP, GTP and acetyl-coA. Fusion is also linked to budding, which is why the term budding and fusing arises.
Vesicles in receptor downregulation
Membrane proteins serving as receptors are sometimes tagged for downregulation by the attachment of ubiquitin. After arriving an endosome via the pathway described above, vesicles begin to form inside the endosome, taking with them the membrane proteins meant for degradation; When the endosome either matures to become a lysosome or is united with one, the vesicles are completely degraded. Without this mechanism, only the extracellular part of the membrane proteins would reach the lumen of the lysosome and only this part would be degraded.
It is because of these vesicles that the endosome is sometimes known as a multivesicular body. The pathway to their formation is not completely understood; unlike the other vesicles described above, the outer surface of the vesicles is not in contact with the cytosol.
Isolated vesicles
Producing membrane vesicles is one of the methods to investigate various membranes of the cell. After the living tissue is crushed into suspension, various membranes form tiny closed bubbles. Big fragments of the crushed cells can be discarded by low-speed centrifugation and later the fraction of the known origin (plasmalemma, tonoplast, etc.) can be isolated by precise high-speed centrifugation in the density gradient. Using osmotic shock, it is possible temporarily open vesicles (filling them with the required solution) and then centrifugate down again and resuspend in a different solution. Applying ionophores like valinomycin can create electrochemical gradients comparable to the gradients inside living cells.
Vesicles are mainly used in two types of research:
Artificial vesicles
Phospholipid vesicles have also been studied in biochemistry. For such studies, a homogeneous phospholipid vesicle suspension can be prepared by extrusion or sonication, injection of a phospholipid solution into the aqueous buffer solution membranes. In this way aqueous vesicle solutions can be prepared of different phospholipid composition, as well as different sizes of vesicles.