
Symbol PQ-loop Pfam clan PQ-loop SMART SM00679 | Pfam PF04193 InterPro IPR006603 TCDB 2.A.123 | |
 | ||
The SWEET family, also known as the PQ-loop, Saliva or MtN3 family (TC# 2.A.123), is a family of sugar transporters and a member of the TOG superfamily. The proteins of the SWEET family have been found in plants, animals, protozoans, and bacteria. Eukaryotic family members have 7 transmembrane segments (TMSs) in a 3+1+3 repeat arrangement.
Contents
Function
Sugar efflux transporters are essential for the maintenance of animal blood glucose levels, plant nectar production, and plant seed and pollen development. Proteins of the SWEET family appear to catalyze facilitated diffusion (entry or export) of sugars across the plant plasma membrane or the endoplasmic reticulum membrane.
Transport Reaction
The generalized reaction catalyzed by known proteins of this family is:
sugars (in) ⇌ sugars (out)Discovery
SWEETs were originally identified in Arabidopsis thaliana, in a screen for novel facilitators of transmembrane glucose transport. In this experiment, several previously uncharacterized membrane proteins were selected to be screened. These uncharacterized membrane proteins were assayed for glucose transport ability by expression in HEK293T (human embryonic kidney) cells, which have negligible glucose transport ability in the normal state. These membrane proteins were co-expressed with a fluorescent FRET (Förster resonance energy transfer) glucose sensor localized to the endoplasmic reticulum (ER). Glucose movement from the cytoplasm to the ER of the HEK293T cells was monitored by quantifying changes in FRET ratio. By using this assay, the first member of the SWEET family, AtSWEET1, was identified. Other potential family members were identified by sequence homology.
Homologues
Chen et al. (2010) reviewed evidence for a new class of sugar transporters, named SWEETs. Those that mediate glucose transport include at least six out of seventeen sugar homologues in Arabidopsis (i.e., TC#s 2.A.123.1.3, 2.A.123.1.5, 2.A.123.1.9, 2.A.123.1.13), two out of over twenty porters in rice (TC#s 2.A.123.1.6 and 2.A.123.1.18), two out of seven homologues in Caenorhabditis elegans (i.e.,TC# 2.A.123.1.10) and the single copy human protein (SLC50A1 of Homo sapiens, TC# 2.A.123.1.4). Arabidopsis SWEET8 (TC# 2.A.123.1.5) is essential for pollen viability. The corn homolog ZmSWEET4c was shown to be involved in seed filling. The rice homologues SWEET11 (TC# 2.A.123.1.13) and SWEET14 are specifically exploited by bacterial pathogens for virulence by means of direct binding of a bacterial effector to the SWEET promoter. Bacterial symbionts and fungal and bacterial pathogens induce the expression of different SWEET genes, indicating that the sugar efflux function of SWEET transporters is targeted by pathogens and symbionts for nutritional gain. The metazoan homologues may be involved in sugar efflux from intestinal, liver, epididymis and mammary cells.
Currently classified members of the SWEET transporter family can be found in the Transporter Classification Database.
SWEETs in plants
Plant SWEETs fall into four subclades. The tomato genome encodes 29 SWEETs. Feng et al. (2015) analyzed the structures, conserved domains, and phylogenetic relationships of these proteins, and also analyzed the transcript levels of SWEET genes in various tissues, organs, and developmental stages in response to exogenous sugar and adverse environmental stress (high and low temperatures).
Plant SWEETs play crucial roles in cellular sugar efflux processes: phloem loading, pollen nutrition and nectar secretion. Plants transport fixed carbon predominantly as sucrose, which is produced in mesophyll cells and imported into phloem cells for translocation throughout the plant. It had not been known how sucrose migrates from sites of synthesis in the mesophyll to the phloem, or which cells mediate efflux into the apoplasm as a prerequisite for phloem loading by the SUT (sucrose transporter family) sucrose-H+ cotransporters. Using optical sucrose sensors, Chen et al. (2012) identified a subfamily of SWEET sucrose efflux transporters. AtSWEET11 (TC# 2.A.123.1.13) and AtSWEET12 localize to the plasma membrane of the phloem. Mutant plants carrying insertions in AtSWEET11 and -12 are defective in phloem loading, thus revealing a two-step mechanism of SWEET-mediated export from parenchyma cells feeding H+-coupled import into the sieve element-companion cell complex. Restriction of intercellular transport to the interface of adjacent phloem cells may be an effective mechanism to limit the availability of photosynthetic carbon in the leaf apoplasm in order to prevent pathogen infections.
SWEET9 in Nectar Secretion
Lin et al., 2014, examined the role of SWEET9 in nectaries. SWEET9 is a member of clade 3. A homologue in petunias had been shown to have an inverse correlation between expression and starch content in nectaries. Mutation and overexpression of SWEET9 in Arabidopsis led to corresponding loss of and increase in nectar secretion, respectively. After showing that SWEET9 is involved in nectar secretion, the next step was to determine at which phase of the process SWEET9 has its function. The 3 options were: phloem unloading, or uptake or efflux from nectary parenchyma. A combination of localization studies and starch accumulation assays showed that SWEET9 is involved in sucrose efflux from the nectary parenchyma.
SWEETs 11, 12, and 15 in Embryo Nutrition
Chen et al., 2015, asked what SWEETs are involved in providing nutrition to an embryo. The team noticed that mRNA and protein for SWEETs 11, 12, and 15 are each expressed at high levels during some stage of embryo development. Each gene was subsequently mutated to generate a sweet11;12;15 triple mutant which lacked activity in each of the three genes. This triple mutant was shown to have delayed embryo development; that is, the seeds of the triple mutant were significantly smaller than that of the wild type at the same time during development. The starch content of the seed coat was higher than the wild-type, and the starch content of the embryo was lower than the wild-type. Additionally, protein levels were shown to be maternally controlled: in a sweet11;12;15 mutant crossed with a wild-type plant, the mutant phenotype was only seen when sweet11;12;15 was used as the maternal plant.
Structure
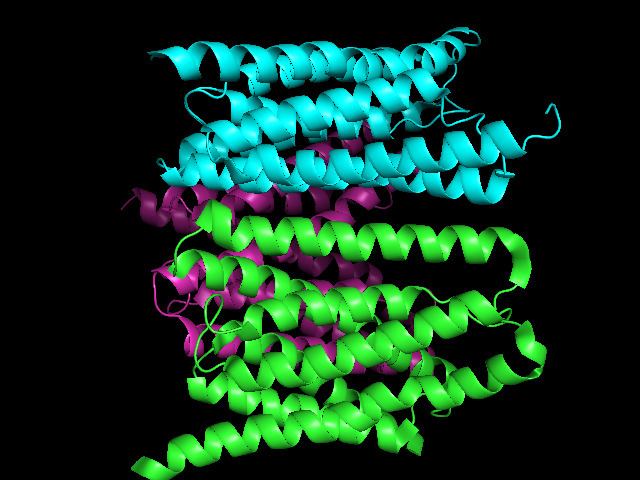
Many bacterial homologues have only 3 TMSs and are half sized, but they nevertheless are members of the SWEET family with a single 3 TMS repeat unit. Other bacterial homologues have 7 TMSs as do most eukaryotic proteins in this family. The SWEET family is large and diverse. Based on 3-D structural analyses, it is likely that these paired 3 TMS SWEET family members function as carriers.
Bacterial SemiSWEETs, consist of a triple-helix bundle in a 1-3-2 conformation, with TM3 sandwiched between TM1 and TM2. SemiSWEETs form semi-symmetrical, parallel dimers, thereby generating the translocation pathway. Two SemiSWEET isoforms have been crystallized, one in an apparently open state and one in an occluded state, indicating that SemiSWEETs and SWEETs are transporters that undergo rocking-type movements during the transport cycle. The structures also show tryptophan and asparagine residues interacting with the sugar; point mutations of these residues to alanine destroys the hexose transport function of SemiSWEET. In SemiSWEETs and SWEETs with more than 3 TM domains, two triple-helix bundles are arranged in a parallel configuration to produce the 6- and (3 + 1 + 3)-transmembrane-helix pores, respectively. Given the similarity of SemiSWEETs and SWEETs to PQ-loop amino acid transporters and to mitochondrial pyruvate carriers (MPCs), the structures characterized by Xu et al., 2014 may also be relevant to other transporters in the SWEET clan. The SWEET family is a member of the TOG superfamily which is believed to have arisen via the pathway:
2 TMSs --> 4 TMSs --> 8 TMSs --> 7 TMSs --> 3 + 3 TMSs.
Several crystal structures are available on RCSB for members of the SWEET/SemiSWEET/PQ-loop/Saliva/MtN3 family.
Oligomerization
Arabidopsis SWEETs homo- and heterooligomerize. Xuan et al., (2013) examined mutant SWEET variants for negative dominance to test if oligomerization is necessary for function. Mutation of the conserved Y57 or G58 residues in SWEET1 (TC# 2.A.123.1.9) led to loss of activity. Coexpression of the defective mutants with functional A. thaliana SWEET1 inhibited glucose transport, indicating that homooligomerization is necessary for function. Collectively, these data imply that the basic unit of SWEETs is a 3-TM unit and that a functional transporter contains at least four such domains. Tao et al., 2015, solved the structure of rice SWEET2 and showed that it forms a homotrimer.