
 | ||
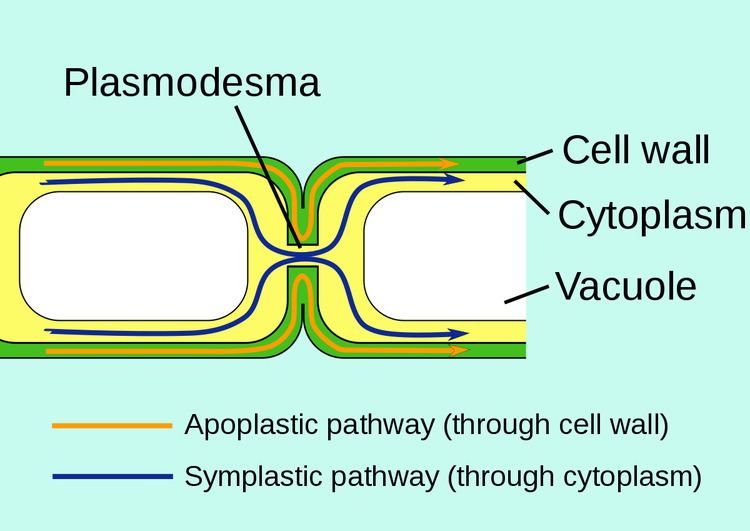
Plasmodesmata (singular: plasmodesma) are microscopic channels which traverse the cell walls of plant cells and some algal cells, enabling transport and communication between them. Plasmodesmata evolved independently in several lineages, and species that have these structures include members of the Charophyceae, Charales, Coleochaetales and Phaeophyceae (which are all algae), as well as all embryophytes, better known as land plants. Unlike animal cells, almost every plant cell is surrounded by a polysaccharide cell wall. Neighbouring plant cells are therefore separated by a pair of cell walls and the intervening middle lamella, forming an extracellular domain known as the apoplast. Although cell walls are permeable to small soluble proteins and other solutes, plasmodesmata enable direct, regulated, symplastic transport of substances between cells. There are two forms of plasmodesmata: primary plasmodesmata, which are formed during cell division, and secondary plasmodesmata, which can form between mature cells.
Contents
Similar structures, called gap junctions and membrane nanotubes, interconnect animal cells and stromules form between plastids in plant cells.
Formation
Primary plasmodesmata are formed when portions of the endoplasmic reticulum are trapped across the middle lamella as new cell wall is laid down between two newly divided plant cells and these eventually become the cytoplasmic connections between cells. Here the wall is not thickened further, and depressions or thin areas known as pits are formed in the walls. Pits normally pair up between adjacent cells. Plasmodesmata can also be inserted into existing cell walls between non-dividing cells (secondary plasmodesmata)
Plasmodesmatal plasma membrane
A typical plant cell may have between 103 and 105 plasmodesmata connecting it with adjacent cells equating to between 1 and 10 per µm2. Plasmodesmata are approximately 50-60 nm in diameter at the midpoint and are constructed of three main layers, the plasma membrane, the cytoplasmic sleeve, and the desmotubule. They can transverse cell walls that are up to 90 nm thick.
The plasma membrane portion of the plasmodesma is a continuous extension of the cell membrane or plasmalemma and has a similar phospholipid bilayer structure.
Cytoplasmic sleeve
The cytoplasmic sleeve is a fluid-filled space enclosed by the plasmalemma and is a continuous extension of the cytosol. Trafficking of molecules and ions through plasmodesmata occurs through this space. Smaller molecules (e.g. sugars and amino acids) and ions can easily pass through plasmodesmata by diffusion without the need for additional chemical energy. Larger molecules, including proteins (for example Green fluorescent protein) and RNA, can also pass through the cytoplasmic sleeve diffusively. Plasmodesmatal transport of some larger molecules is facilitated by mechanisms that are currently unknown. One mechanism of regulation of the permeability of plasmodesmata is the accumulation of the polysaccharide callose around the neck region to form a collar, thereby reducing the diameter of the pore available for transport of substances.
Desmotubule
The desmotubule is a tube of appressed (flattened) endoplasmic reticulum that runs between two adjacent cells. Some molecules are known to be transported through this channel, but it is not thought to be the main route for plasmodesmatal transport.
Around the desmotubule and the plasma membrane areas of an electron dense material have been seen, often joined together by spoke-like structures that seem to split the plasmodesma into smaller channels. These structures may be composed of myosin and actin, which are part of the cell's cytoskeleton. If this is the case these proteins could be used in the selective transport of large molecules between the two cells.
Transport
Plasmodesmata have been shown to transport proteins (including transcription factors), short interfering RNA, messenger RNA, viroids, and viral genomes from cell to cell. One example of a viral movement proteins is the tobacco mosaic virus MP-30. MP-30 is thought to bind to the virus's own genome and shuttle it from infected cells to uninfected cells through plasmodesmata. Flowering Locus T protein moves from leaves to the shoot apical meristem through plasmodesmata to initiate flowering.
Plasmodesmata are also used by cells in phloem, and symplastic transport is used to regulate the sieve-tube cells by the companion cells.
The size of molecules that can pass through plasmodesmata is determined by the size exclusion limit. This limit is highly variable and is subject to active modification. For example, MP-30 is able to increase the size exclusion limit from 700 Daltons to 9400 Daltons thereby aiding its movement through a plant. Also, increasing calcium concentrations in the cytoplasm, either by injection or by cold-induction, has been shown to constrict the opening of surrounding plasmodesmata and limit transport.
Several models for possible active transport through plasmodesmata exist. It has been suggested that such transport is mediated by interactions with proteins localized on the desmotubule, and/or by chaperones partially unfolding proteins, allowing them to fit through the narrow passage. A similar mechanism may be involved in transporting viral nucleic acids through the plasmodesmata.