
Organism Arthrospira maxima Alt. symbols slr1963 UniProt P83689 | Symbol Ocp PDB 1M98 | |
 | ||
Orange carotenoid protein (OCP) is a water-soluble protein which plays a role in photoprotection in diverse cyanobacteria. It is the only photoactive protein known to use a carotenoid as the photoresponsive chromophore. The protein consists of two domains, with a single keto-carotenoid molecule non-covalently bound between the two domains. It is a very efficient quencher of excitation energy absorbed by the primary light-harvesting antenna complexes of cyanobacteria, the phycobilisomes. The quenching is induced by blue-green light. It is also capable of preventing oxidative damage by directly scavenging singlet oxygen (1O2).
Contents
History
OCP was first described in 1981 by Holt and Krogmann who isolated it from the unicellular cyanobacterium Arthrospira maxima, although its function would remain obscure until 2006. The crystal structure of the OCP was reported in 2003 and the protein was shown to be an effective quencher of singlet oxygen. In 2000, it was demonstrated that cyanobacteria could perform photoprotective fluorescence quenching independent of lipid phase transitions, differential transmembrane pH, and inhibitors. The action spectrum for this quenching process suggested the involvement of carotenoids, and the specific involvement of the OCP was later demonstrated by Kirilovsky and coworkers in 2006. In 2008, OCP was shown to require photoactivation by strong blue-green light for its photoprotective quenching function.
Physiological significance
For a long time, cyanobacteria were considered incapable of performing non-photochemical quenching (NPQ) as a photoprotective mechanism, relying instead on a mechanism of energy redistribution between the two photosynthetic reaction centers, PSII and PSI, known as "state transitions".
OCP is found in a majority of cyanobacterial genomes, with remarkable conservation of its amino acid sequence, implying evolutionary constraints to preserve an important function. Mutant cells engineered to lack OCP photobleach under high light and become photoinhibited more rapidly under fluctuating light. Under nutrient stress conditions, which are expected to be norm in marine environments, photoprotective mechanisms such as OCP become important even at lower irradiances.
This protein is not found in chloroplasts, and appears to be specific to cyanobacteria.
Photoactivity
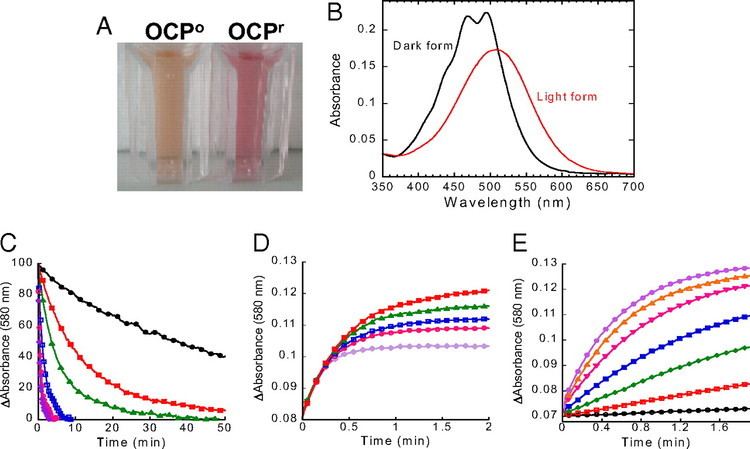
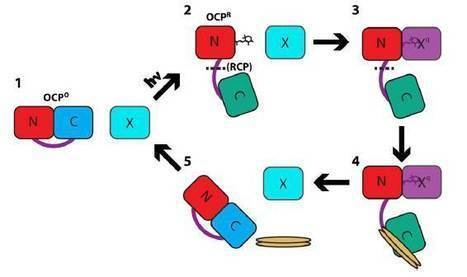
Upon illumination with blue-green light, OCP switches from an orange form (OCPO) to a red form (OCPR). The reversion of OCPR to OCPO is light independent and occurs slowly in darkness. OCPO is considered the dark, stable form of the protein, and does not contribute to phycobilisome quenching. OCPR is considered to be essential for induction of the photoprotection mechanism. The photoconversion from the orange to red form has a poor light efficiency (very low quantum yield), which helps to ensure the protein's photoprotective role only functions during high light conditions; otherwise, the dissipative NPQ process could unproductively divert light energy away from photosynthesis under light-limiting conditions.
Energy quenching
As evidenced by a decreased fluorescence, OCP in its red form is capable of dissipating absorbed light energy from the phycobilisome antenna complex. According to Rakhimberdieva and coworkers, about 30-40% of the energy absorbed by phycobilisomes does not reach the reaction centers when the carotenoid-induced NPQ is active. The exact mechanism and quenching site in both the carotenoid as well as the phycobilisome still remain uncertain. The linker polypeptide ApcE in the allophycocyanin (APC) core of the phycobilisomes is known to be important, but is not the site of quenching. Several lines of evidence suggest that it is the 660 nm fluorescence emission band of the APC core which is quenched by OCPR. The temperature dependence of the rate of fluorescence quenching is similar to that of soluble protein folding, supporting the hypothesis that OCPO slightly unfolds when it converts to OCPR.
Singlet oxygen quenching
Contributed by the auxiliary function of carotenoids as quenchers of singlet oxygen, a photoprotective role of OCP has also been demonstrated under strong orange-red light, which are conditions where OCP cannot be photoactivated for its energy-quenching role. This is significant because all oxygenic phototrophs have a particular risk of oxidative damage initiated by singlet oxygen (1O2), which is produced when their own light-harvesting pigments act as photosensitizers.
3D structure
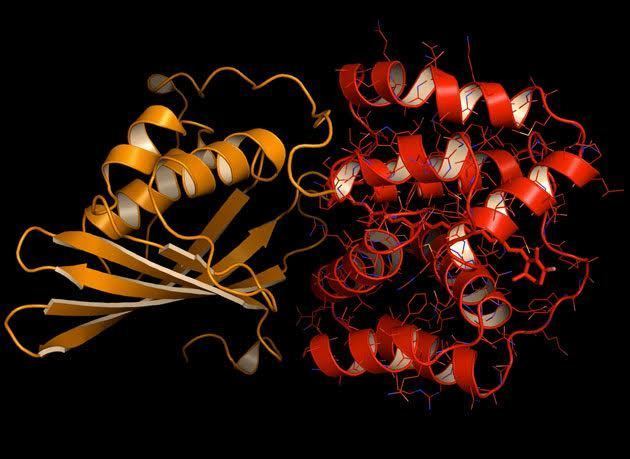
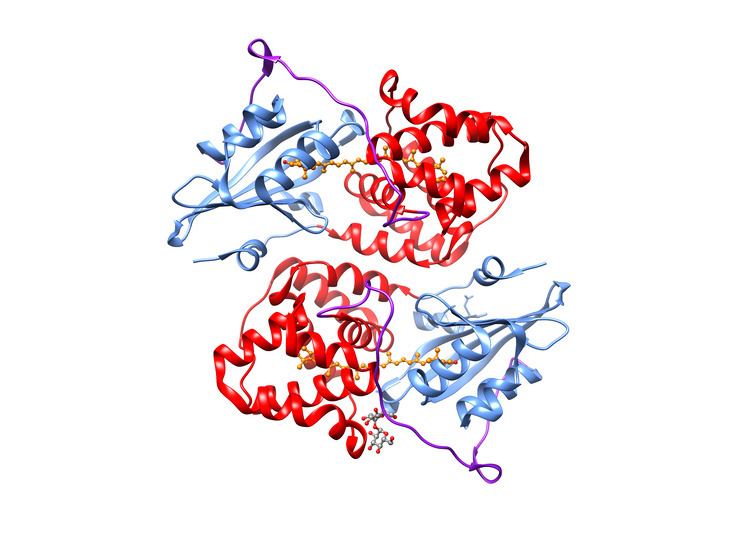
The three-dimensional protein structure of OCP (in the OCPO form) was solved in 2003, before its photoprotective role had been elucidated. The 35 kDa protein contains two structural domains: an all-α-helical N-terminal domain (NTD) consisting of two interleaved 4-helix bundles, and a mixed α/β C-terminal domain (CTD). The two domains are connected by an extended linker. In OCPO, the carotenoid spans both domains, which are tightly associated in this form of protein.
Protein–protein interactions
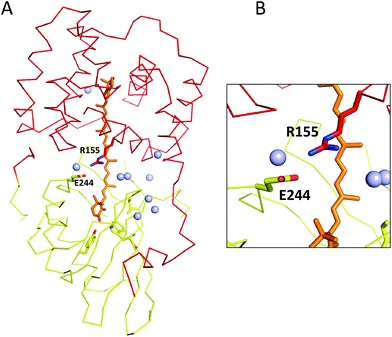
The OCP participates in key protein–protein interactions that are critical to its photoprotective function. The activated OCPR form binds to allophycocyanin in the core of the phycobilisome and initiates the OCP-dependent photoprotective quenching mechanism. Another protein, the fluorescence recovery protein (FRP), interacts with the CTD in OCPR and catalyzes the reaction which reverts it back to the OCPO form. Because OCPO cannot bind to the phycobilisome antenna, FRP effectively can detach OCP from the antenna and restore full light-harvesting capacity.
Evolution
The primary structure (amino acid sequence) is highly conserved among OCP sequences, and the full-length protein is usually co-located on the chromosome with the FRP at an adjacent locus. Often, biosynthetic genes for ketocarotenoid synthesis (e.g., CrtW) are nearby. These conserved functional linkages underscore the evolutionary importance of the OCP style of photoprotection for many cyanobacteria.
There is also a variety of evolutionarily related genes which encode proteins with only one of the two domains present in OCP. The N-terminal domain (NTD), "Carot_N", is found only in cyanobacteria, but exhibits a considerable amount of gene duplication. The C-terminal domain (CTD), however, is homologous with the widespread NTF2 superfamily, which shares a protein fold with its namesake, nuclear transport factor 2, as well as around 20 other subfamilies of proteins with functions as diverse as limonene-1,2-epoxide hydrolase, SnoaL polyketide cyclase, and delta-5-3-ketosteroid isomerase (KSI). Most, if not all, of the members of the NTF2 superfamily form oligomers, often using the surface of their beta sheet to interact with another monomer or other protein.
Applications
Its water-solubility, together with its status as the only known photoactive protein containing a carotenoid, makes the OCP a valuable model for studying solution-state energetic and photophysical properties of carotenoids, which are a diverse class of molecules found across all domains of life. Moreover, carotenoids are widely investigated for their properties as anti-oxidants, and thus the protein may serve as a template for delivery of carotenoids for therapeutic purposes in human medicine.
Because of its high efficiency of fluorescence quenching, coupled to its low quantum yield of photoactivation by specific wavelengths of light, OCP has ideal properties as a photoswitch and has been proposed as a novel system for developing optogenetics technologies and may have other applications in optofluidics and biophotonics.