
Rank Class | Scientific name Monogenea Higher classification Flatworm | |
 | ||
Similar | ||
Intro to the monogeneans part i
Monogenea (adj. monogenean) are a group of largely ectoparasitic members of the flatworm phylum Platyhelminthes, class Monogenea.
Contents
Dactylogyrids a gill monogenean parasite of fishes
Characteristics
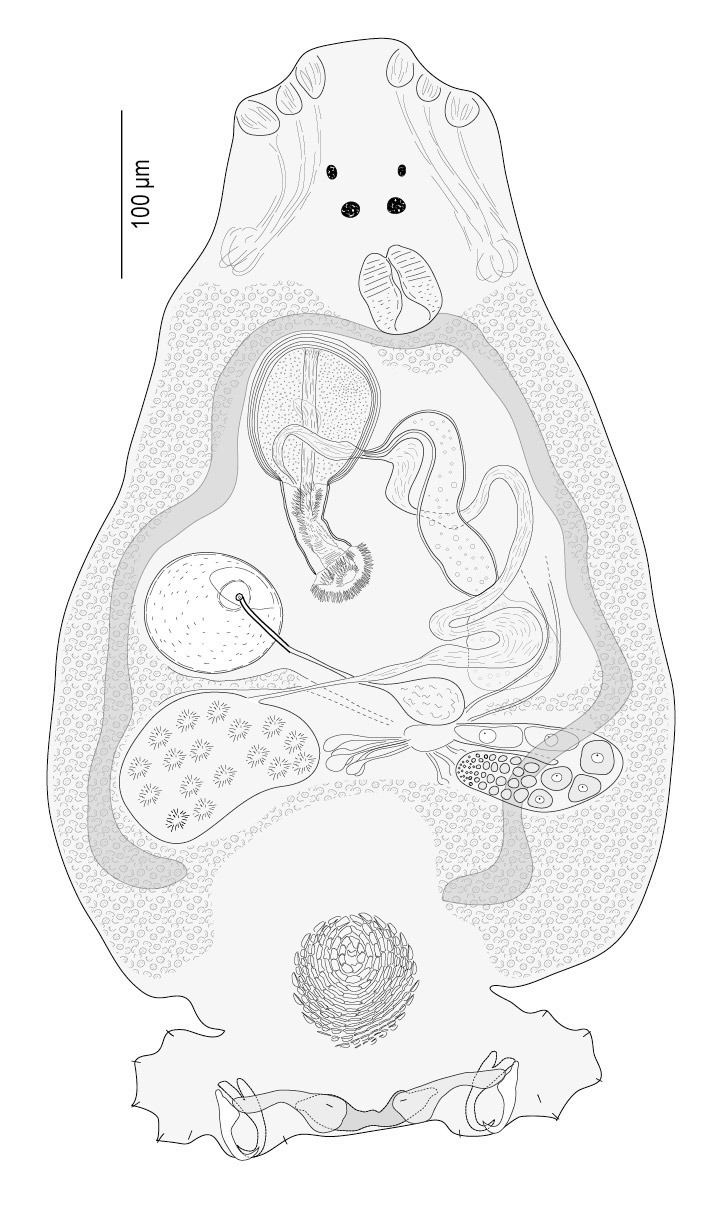
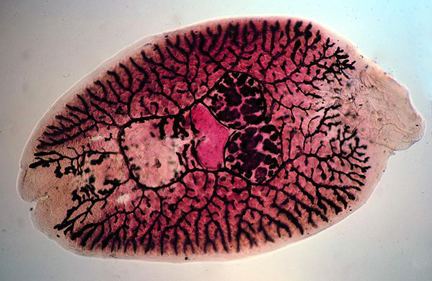
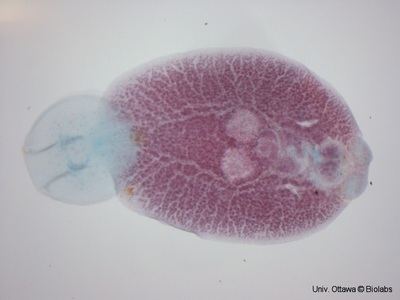
Monogenea are small parasitic flatworms mainly found on skin or gills of fish. They are rarely longer than about 2 cm. A few species infecting certain marine fish are larger and marine forms are generally larger than those found on fresh water hosts. Monogeneans lack respiratory, skeletal and circulatory systems and have no or weakly developed oral suckers. Monogenea attach to hosts using hooks, clamps and a variety of other specialized structures. They are often capable of dramatically elongating and shortening as they move. Biologists need to ensure that specimens are completely relaxed before measurements are taken.
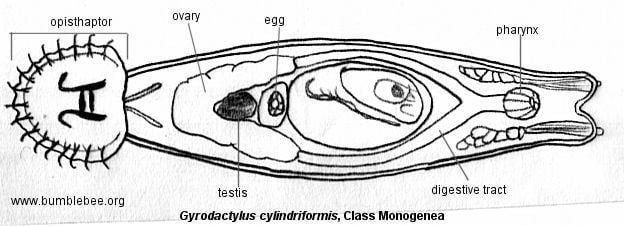
Like all ectoparasites, monogeneans have well-developed attachment structures. The anterior structures are collectively termed the prohaptor, while the posterior ones are collectively termed the opisthaptor, or simply haptor. The posterior opisthaptor with its hooks, anchors, clamps etc. is typically the major attachment organ.
Like other flatworms, Monogenea have no true body cavity (coelom). They have a simple digestive system consisting of a mouth opening with a muscular pharynx and an intestine with no terminal opening (anus). Generally, they also are hermaphroditic with functional reproductive organs of both sexes occurring in one individual. Most species are oviparous but a few are viviparous. Monogenea are Platyhelminthes and therefore are among the lowest invertebrates to possess three embryonic germ layers—endoderm, mesoderm, and ectoderm. In addition, they have a head region that contains concentrated sense organs and nervous tissue (brain).
Systematics and evolution
The ancestors of Monogenea were probably free-living flatworms similar to modern Turbellaria. According to the more widely accepted view, "rhabdocoel turbellarians gave rise to monogeneans; these, in turn, gave rise to digeneans, from which the cestodes were derived. Another view is that the rhabdocoel ancestor gave rise to two lines; one gave rise to monogeneans, who gave rise to digeneans, and the other line gave rise to cestodes" [1] .
There are about 50 families and thousands of described species.
Some parasitologists divide Monogenea into two (or three) subclasses based on the complexity of their haptor: Monopisthocotylea have one main part to the haptor, often with hooks or a large attachment disc, whereas Polyopisthocotylea have multiple parts to the haptor, typically clamps. These groups are also known as Polyonchoinea and Heteronchoinea, respectively. Polyopisthocotyleans are almost exclusively gill-dwelling blood feeders, whereas Monopisthocotyleans may live on the gills, skin and fins.
Monopisthocotylea include:
All of which can cause epizootics in freshwater fish when raised in aquaculture.
Polyopisthocotylea include:
Ecology and life cycle
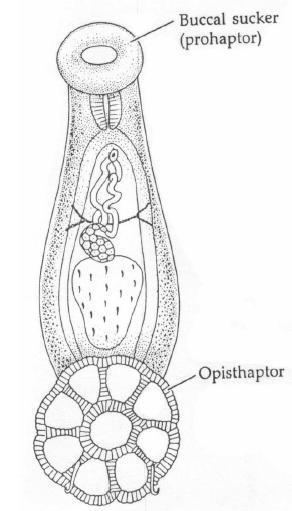
Monogeneans possess the simplest life cycle among the parasitic platyhelminths. They have no intermediate hosts and are ectoparasitic on fish (seldom in the urinary bladder and rectum of cold-blooded vertebrates). Although they are hermaphrodites, the male reproductive system becomes functional before the female part. The eggs hatch releasing a heavily ciliated larval stage known as an oncomiracidium. The oncomiracidium has numerous posterior hooks and is generally the life stage responsible for transmission from host to host.
No known monogeneans infect birds, but one (Oculotrema hippopotami) infects mammals, parasitizing the eye of the hippopotamus.