
 | ||
Membrane curvature is the geometrical measure or characterization of the curvature of membranes. The membranes can be naturally occurring or man-made (synthetic). An example of naturally occurring membrane is the lipid bilayer of cells, also known as cellular membranes. Synthetic membranes can be obtained by preparing aqueous solutions of certain lipids. The lipids will then "aggregate" and form various phases and structures. According to the conditions (concentration, temperature, ionic strength of solution, etc.) and the chemical structures of the lipid, different phases will be observed. For instance, the lipid POPC (palmitoyl oleyl phosphatidyl choline) tends to form lamellar vesicles in solution, whereas smaller lipids (lipids with shorter acyl chains, up to 8 carbons in length), such as detergents, will form micelles if the CMC (critical micelle concentration) is reached.
Contents
Basic Geometry of Curvature
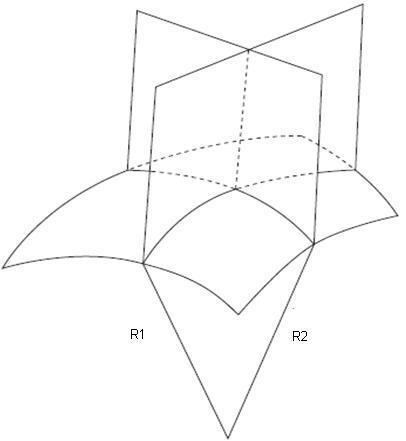
A biological membrane is commonly described as a two-dimensional surface, which spans a three-dimensional space. So, to describe membrane shape, it is not sufficient to determine the membrane curling that is seen in a single cross-section of the object, because in general there are two curvatures that characterize the shape each point in space. Mathematically, these two curvatures are called the principal curvatures, c1 and c2, and their meaning can be understood by the following thought experiment. If you cross-section the membrane surface at a point under consideration using two planes that are perpendicular to the surface and oriented in two special directions called the principal directions, the principal curvatures are the curvatures of the two lines of intercepts between the planes and the surface which have almost circular shapes in close proximity to the point under consideration. The radii of these two circular fragments, R1 and R2, are called the principal radii of curvature, and their inverse values are referred to as the two principal curvatures.
The principal curvatures C1 and C2 can vary arbitrarily and thereby give origin to different geometrical shapes, such as cylinder, plane, sphere and saddle. Analysis of the principal curvature is important, since a number of biological membranes possess shapes that are analogous to these common geometry staples. For instance, prokaryotic cells such as cocci, rods, and spirochette display the shape of a sphere, and the latter two the shape of a cylinder. Erythrocytes, commonly referred to as red blood cells, have the shape of a saddle, although these cells are capable of some shape deformation. The table below lists common geometric shapes and a qualitative analysis of their two principal curvatures.
Even though often membrane curvature is thought to be a completely spontaneous process, thermodynamically speaking there must be factors actuating as the driving force for curvature to exist. Currently, there are some postulated mechanisms for accepted theories on curvature; nonetheless, undoubtedly two of the major driving forces are lipid composition and proteins embedded and/or bound to membranes.
Lipid Spontaneous Curvature
Perhaps the most simple and intuitive driving force in membrane curvature is the natural spontaneous curvature exhibited by some lipids. This is because, depending on their chemical structures, lipids tend to curve with a slight spontaneously negative or positive curvature. Lipids such as DOPC (dioleoyl phosphatidyl choline), diacyl glycerol, dioleyl phosphatidylethanolamine (DOPE) and cholesterol exhibit a negative spontaneous curvature. On the other hand, lipids with smaller acyl chain area to polar head group area ratio tend to curve positively, in other words they exhibit positive spontaneous curvature. The table below lists experimentally determined spontaneous curvatures for different lipids in DOPE (dioleyl phosphatidyl ethanolamine).
The energy requirements to generate a cylinder shaped cell from an originally flat membrane can be expressed as
FCyl =πxLxKb(1/R - 2JB) where L is the length of the cylinder, JB is the difference between the spontaneous curvature, Js, for the lipids in the inner and outer leaflet divided by two, and Kb is the bending modulus of the bilayer.
The radii of membrane cylinders that form in intracellular membrane-transport pathways are typically ~25–30 nm. So, the spontaneous curvature necessary to generate such cylinders equals ~(1/50) nm–1. As JB results from a difference in the spontaneous curvatures of the monolayers, an unusual membrane lipid composition would be required to produce such curvature. The lipids cholesterol, dioleoylphosphatidylethanolamine(DOPE) and diacylglycerol are characterized by strongly negative spontaneous curvatures (figure 1) and therefore have the potential to generate a large membrane curvature. However, even for these lipids, the required JB can be reached only if they are extensively concentrated in the internal monolayer.
Proteins can Induce Curvature
Some biologically occurring lipids do exhibit spontaneous curvature which could explain the shapes of biological membranes. Nevertheless, calculations show that spontaneous lipid curvature alone is either insufficient or would require conditions that are unrealistic to drive the degree of curvature observed in most cells. It is now known that lipid curvature is "aided" by protein structures in order to generate complete cellular curvature. A classical example of such interactions is the activity of the protein clathrin. Clathrin is involved in cellular endocytosis and is sequestrated by specific signaling molecules. Clathrin can attach to adaptor protein complexes on the cellular membrane, and it polymerizes to drive greater curvature, resulting in endocytosis of a vesicular unit.
Another example of protein interactions that directly affect membrane curvature is that of the BAR (Bin, amphiphysin, Rvs’) domain. The BAR domain is present in a large family of proteins. This domain is rigid, relative to the cellular lipid bilayer, and it exhibits a "banana" shape. Upon binding, the membrane's curvature is increased by the rigid domain.
One more case of protein interaction that induces and/or aids curvature is the class of proteins such as epsin. Epsin has several alpha helices that possess amphipathic properties, which allows it to partition between the hydrophobic core of the membrane and the hydrophilic aqueous environment. Another interesting characteristic of epsin and other proteins that bind to membranes is the fact that it shows high binding affinity for a fairly common membrane lipid, phosphatidylinositol 4,5-bisphosphate (PI-4,5-P2). Unlike other proteins that simply bend the membrane through sheer rigidity, epsin is a globular soluble protein and thus not rigid. The insertion of its helices into the membrane force the neighboring lipids of the leaflet that has been bound to expand laterally. This displacement of lipids on only one of the leaflets increases the bilayer's curvature.
The figure illustrates the different mechanisms through which proteins can aid and/or induce membrane curvature. In A, an illustration of a BAR domain present in a number of proteins. The curvature is induced by the very shape of this proteic region. This domain attaches to the lipid bilayer through strong coulombic interactions. This idea is supported by the existence of positively charged amino acid residues in the concave region of the BAR domain. These amino acids would come into contact with the negatively charged polar head groups of lipids in the bilayer. This form phenomenon is also referred to as the "scaffold mechanism".
B shows a protein coating that induces curvature. As mentioned above, proteins such as clathrin are recruited to the membrane through signaling molecules and assemble into larger polymeric structures that form a rigid structure which serves as a frame for the membrane. Clathrin binds to its receptors that are present in the membrane.
C illustrates a slightly different mechanism. In this case, the membrane-bending protein does not exhibit intrinsic rigidity. Instead they are often globular and soluble. The protein epsin is an example. Epsin has an ENTH (epsin N-terminal homology) domain which inserts its amphipathic alpha helix into the membrane. Epsin has high binding affinity for the membrane if PI-4,5-P2 is present.