
 | ||
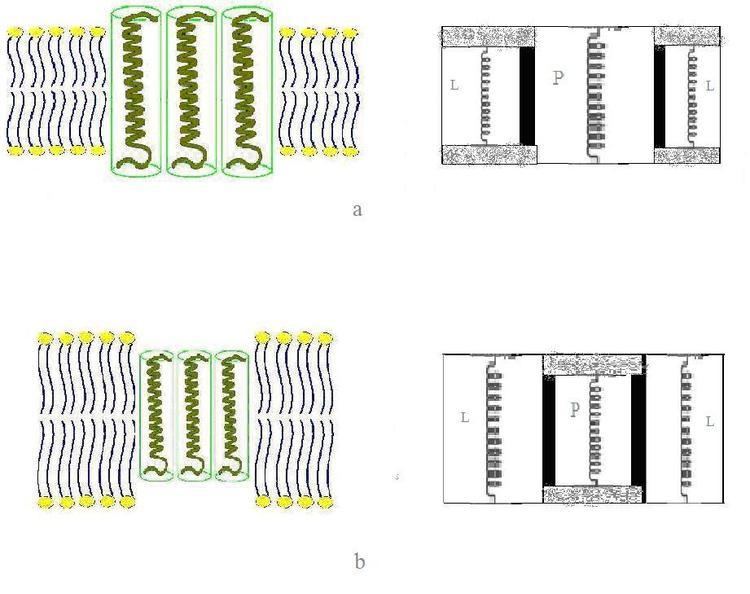
Hydrophobic mismatch is the difference between the hydrophobic length of the integral proteins and the hydrophobic thickness of the membrane they span. Under the consideration of energy requirement, in order to avoid unfavorable exposure of hydrophobic surfaces to a hydrophilic environment, the hydrophobic length of the integral proteins is supposed to be approximately equal to the hydrophobic bilayer thickness.
Contents
Structure and organization
A biological membrane is a kind of amphipathic layer that acts as a barrier within or around a cell. In most case, it is a lipid bilayer, composed of a double layer of lipid molecules and proteins that may constitute close to 50% of membrane content. The interaction of integral proteins with the lipids inside membrane bilayer is of great importance for membrane function.
The thickness of biological membrane is approximately same with the hydrophobic core of lipid bilayer. At the same time, the protein embedded inside one membrane could have different hydrophobic length, for example Escherichia coli, a Gram negative bacterium that is commonly found in the lower intestine of warm-blooded animals, the predicted length of inner membrane enzyme leader peptidase for the second transmembrane helix is only 15 amino acids long. While resides in the same membrane, the helices of lactose permease has the mean length of 24±4 residues long. Vice versa, membrane proteins with the same length can be encountered in bilayers of different thickness. Like in eukaryotic cells, due to their higher content of cholesterol and sphingomyelin, the plasma membrane is much thicker than the membranes of the endoplasmic reticulum. Yet all proteins that are abundant in the plasma membrane are initially integrated in the endoplasmic reticulum upon synthesis on ribosomes.
Possible adaptations to mismatch
In order to avoid unfavorable exposure of hydrophobic surfaces to a hydrophilic environment, biological membrane tends to make some adaptations to such mismatch. In various other systems, is that an integral protein tends to surround itself by lipids of matching size and shape. Since proteins are relatively rigid, whereas lipid hydrocarbon chains are flexible, the condition of hydrophobic matching can be fulfilled by stretching, squashing, and/or tilting of the lipid chains
Protein aggregation
Since Mouritsen and Bloom proposed the detailed thermodynamic model, which includes adaptation of the lipids and induction of protein segregation at a more extreme mismatch in their “Mattress Model”, more additional insight into mismatch-induced protein aggregation has been obtained. Also some experimental evidence that a hydrophobic mismatch can lead to protein aggregation in fluid bilayer were founded. Electron microscopy studies on bacteriorhodopsin, reconstituted in saturated and unsaturated fluid PC bilayers with varying chain length, showed that protein aggregation occurred only with a rather large mismatch, and that bilayer thicknesses of 4 angstrom thicker and 10 angstrom thinner than the estimated hydrophobic length of the protein are allowed without induction of significant aggregation.
Helix tilt
Tilt is also a possible result if the hydrophobic part of a peptide or protein is too long to span the membrane. A previous study on lactose permease of E. coli showed that upon reconstitution of the protein in PE/PG (3/1) lipid bilayer, an increase in helix tilt occurs at increasing protein content. This tilt was accompanied by a decrease in lipid order, which results in a decrease in bilayer thickness, suggesting that it is a mismatch related response. In large proteins that span the membrane multiple times, changes in helical tilt may occur with little effect on lipid packing. However, for a single transmembrane helix, it is possible that a tilt would cause a strain on the surrounding lipids to accommodate the helix in the bilayer. Thus, a large degree of tilting can be a less favorable option for single transmembrane proteins.
Surface orientation
Relatively small hydrophobic peptides may not be able to integrate into the membrane, and in response adopt an orientation at the membrane surface. The experimental evidence was shown by a fluorescence study on an artificial peptide with a 19 amino acid long hydrophobic sequence of mainly leucines and flanked on both sides with lysines as anchoring residues. The results indicated that a conversion from a dominant transmembrane to parallel orientation of the peptide could be induced by modulating bilayer thickness via addition of cholesterol or by increasing lipid chain length.
Backbone conformation change
To obtain detailed information on the consequences of mismatch for the conformation of peptides and proteins in lipid bilayer, small membrane-spanning peptides are most suitable. Still need some studies.
Theories for the mismatch effects
Since the diverse possibility of the hydrophobic mismatch effects, there is a need to understand this puzzling behavior and to isolate the various factors which bring about the net result of hydrophobic mismatch. Many theoretical approaches have been applied to the effects of mismatch. Typically two trails have been dominated, phenomenological trail which based on treating the membrane as an elastic sheet is used mostly. together with few previously. but more and more useful microscopic approach. Some important theory approaches used in the history are shown here:
Mattress model
Mattress model was proposed as a phenomenological theory approach in 1984 by Mouritsen and Bloom. It is a two-component real solution theory based on the theory of nonideal solutions and hence allows for phase separation. In their model, they relate the energy stored in the undulations of the membrane surface caused by the mismatch to the elastic properties of the lipids and proteins. They do not include microscopic detail of the lipids, but use as input the known thermodynamic properties of the pure lipid system. They also include indirect lipid-protein interactions induced by the mismatch as well as direct lipid-protein van der Waals-like interactions between the hydrophobic parts of the lipid bilayer and the proteins. The excess "hydrophobic effect" associated with the lipid-protein hydrophobic mismatch, and the elastic deformation free energy of the lipid chains near the protein. The interaction potentials are estimated based on experimental data derived from thermodynamic and mechanical measurements of membrane properties.
Monte Carlo simulation scheme
The mattress model was later replicated in a Monte Carlo simulation scheme by Sperotto and Mouritsen. They allowed for different microstates of the lipids, classified according to Pink’s 10-state model. hence enabling a pure lipid bilayer phase transition. This version of the model provides a connection between the microscopic characteristics of the system and its thermodynamic behavior.
Molecular theory
A major theoretical advance was the work of Fattal and Ben-Shaul. who provided a molecular theory for the behavior of the lipid chains of the membrane. The peptides, with their hydrophobic length, were treated as providing a boundary condition on the configuration of the lipid chains. This molecular modeling was combined with phenomenological free energy contributions describing lipid head group repulsion and membrane solvent surface tension. Duque et al. described the effects of an embedded protein in a bilayer via molecular theory, which yielded the free energy of the entire system.
Effect of mismatch
The membrane hydrophobic mismatch has the possible effects in more complex biological membranes and possible significance for biological membrane processes, like in the protein sorting, lipid raft.
Protein sorting
In eukaryotic cells, the level of cholesterol increases through the secretory pathway, from the endoplasmic reticulum to the Golgi to the plasma membrane, suggesting a concomitant increase in membrane thickness. In line with this, the average length of transmembrane domains of plasma membrane proteins typically is five amino acids longer than the average length of proteins from the Golgi. Experimental evidence was obtained that protein sorting in the Golgi may be based on this length difference: for several proteins that normally reside in the Golgi, it was shown that increasing their hydrophobic length can reroute the proteins to the plasma membrane, or vice versa, that decreasing the hydrophobic length of proteins from the plasma membrane can cause their retention in the Golgi.
Lipid raft
One type of membrane heterogeneity, termed rafts, is enriched in cholesterol, sphingomyelin (SM), and certain membrane proteins. Rafts have putative roles in many physiological processes, such as signal transduction, endocytosis, apoptosis, protein trafficking, and lipid regulation. Raft lipids typically have saturated hydrocarbon chains. Cholesterol, a molecule that is enriched in lipid rafts, has a shorter hydrophobic length. Hence, because of the differences in lipid thickness between raft and nonraft lipids, and since energetically it can be expected that the lengths of the hydrophobic moieties of neighboring membrane components will be approximately equal to avoid unfavorable exposure of hydrophobic surfaces to a hydrophilic environment, it is reasonable to assume that nonraft lipids should coalesce and predominantly give rise to smaller bilayer thicknesses compared to raft lipids.