
Symbol ERM InterPro IPR011259 SUPERFAMILY 1ef1 | Pfam PF00769 SCOP 1ef1 Pfam structures | |
 | ||
The ERM protein family consists of three closely related proteins, ezrin, radixin and moesin. The three paralogs, ezrin, radixin and moesin, are present in vertebrates, whereas other species have only one ERM gene. Therefore, in vertebrates these paralogs likely arose by gene duplication.
Contents
ERM proteins are highly conserved throughout evolution. More than 75% identity is observed in the N-terminal and the C-terminal of vertebrates (ezrin, radixin, moesin), Drosophila (dmoesin) and C. elegans (ERM-1) homologs.
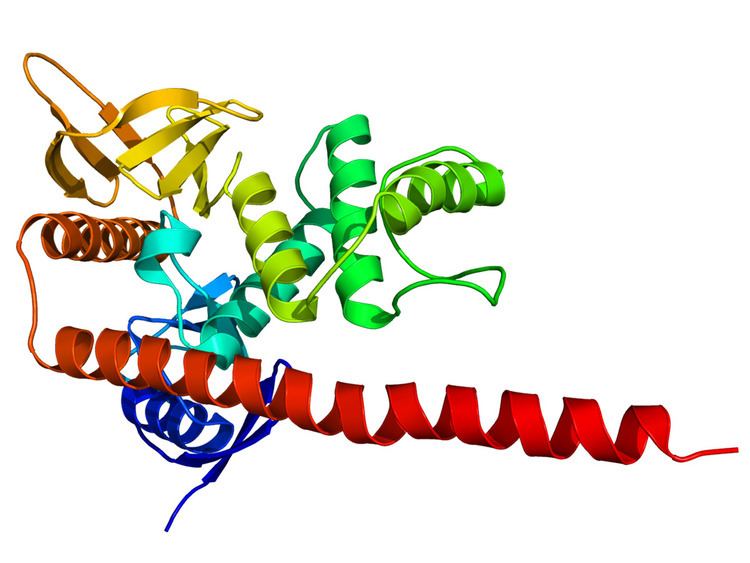
Structure
ERM molecules contain the following three domains:
Ezrin, radixin and merlin also contain a polyproline region between the central helical and C-terminal domains.
Function
ERM proteins crosslink actin filaments with plasma membranes. They co-localize with CD44 at actin filament-plasma membrane interaction sites, associating with CD44 via their N-terminal domains and with actin filaments via their C-terminal domains.
ERM proteins Moesin directly binds to microtubules via its N-terminal FERM domain in vitro and stabilizes microtubules at the cell cortex in vivo. This interaction is required for specific ERM-dependent functions in mitosis.
Activation
ERM proteins are highly regulated proteins. They exist in two forms:
In culture cells, ERM proteins mainly exhibit the folded conformation (about 80-85%).
The current model for ERM proteins activation is a two-steps mechanism: