
 | ||
CRISPR interference (CRISPRi) is a genetic perturbation technique that allows for sequence-specific repression or activation of gene expression in prokaryotic and eukaryotic cells.
Contents
- Background
- Transcriptional regulation
- Repression
- Activation
- Improvements on the efficiency
- SunTag protein scaffold
- Other methods
- Gene knockdown
- Allelic series
- Loci imaging
- Stem cells
- Genetic screening
- Advantages
- Limitations
- References
Based on the bacterial genetic immune system - CRISPR (clustered regularly interspaced short palindromic repeats) pathway, the technique provides a complementary approach to RNA interference. The difference between CRISPRi and RNAi, though, is that CRISPRi regulates gene expression primarily on the transcriptional level, while RNAi controls genes on the mRNA level.
Background
Many bacteria and most archaea have an adaptive immune system which incorporates CRISPR RNA (crRNA) and CRISPR-associated (cas) genes.
The CRISPR interference technique was first reported by researchers at the University of California at San Francisco and University of California at Berkeley in 2013. This minimal CRISPR system has been successfully adapted for gene knockout and gene knockdown in many model organisms (bacteria, yeast, fruit flies, zebrafish, mice,). A complementary technology uses catalytically dead Cas9 (usually denoted dead Cas9 or dCas9) lacking endonuclease activity to regulate genes in an RNA-guided manner. Targeting specificity is determined by complementary base-pairing of a single guide RNA (sgRNA) to the genomic loci. sgRNA is a chimeric noncoding RNA that can be subdivided into three regions: a 20 nt base-pairing sequence, a 42 nt dCas9-binding hairpin and a 40 nt terminator.
When designing a synthetic sgRNA, only the 20 nt base-pairing sequence is modified from the overall template. Additionally, secondary variables must be considered: off-target effects (for which a simple BLAST run of the base-pairing sequence is required), maintenance of the dCas9-binding hairpin structure and to ensure no restriction sites are present in the new sgRNA as this may pose a problem in downstream cloning steps. Due to the simplicity of sgRNA design, this technology is amenable to genome-wide scaling. CRISPRi relies on the generation of catalytically inactive Cas9. This is accomplished by introducing point mutations in the two catalytic residues (D10A and H840A) of the gene encoding Cas9. In doing so, dCas9 is unable to cleave dsDNA but retains the ability to target DNA. Taken together sgRNA and dCas9 provide a minimum system for gene-specific regulation in any organism.
Transcriptional regulation
Although the term CRISPRi was initially coined to describe transcriptional interference for gene silencing, it can also be used to describe activation of transcription and epigenetic modifications.
Repression
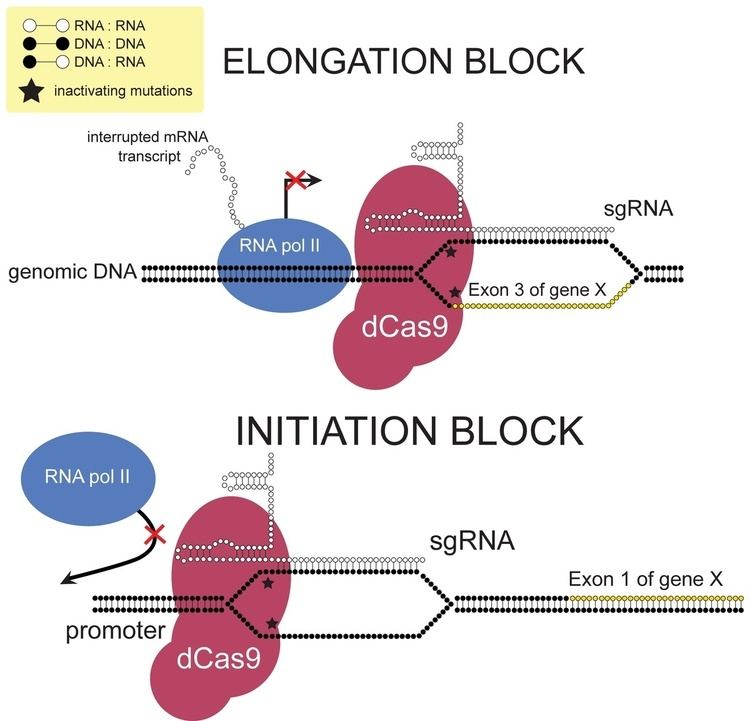
CRISPRi can sterically repress transcription in two ways – by blocking transcriptional initiation or elongation. This is accomplished by designing sgRNA complementary to the promoter or exonic sequences, respectively. The level of transcriptional repression for exonic sequences is strand-specific. sgRNA complementary to the non-template strand more strongly represses transcription compared to sgRNA complementary to the template strand. One hypothesis to explain this effect is from the activity of helicase, which unwinds the RNA:DNA heteroduplex ahead of RNA pol II when the sgRNA is complementary to exons of the template strand. In prokaryotes, this steric inhibition can repress transcription of the target gene by almost 99.9%. Whereas in human cells, up to 90% repression was observed.
CRISPRi can also repress transcription via an effector domain. Fusing a repressor domain to dCas9 allows transcription to be further repressed by inducing heterochromatinization. For example, the well-studied Krüppel associated box (KRAB) domain can be fused to dCas9 to repress transcription of the target gene up to 99% in human cells.
Activation
CRISPRi can be used to activate transcription of the target gene by fusing a transcriptional activator to dCas9. For example, the transcriptional activator VP16 can increase gene expression by up to 25-fold in human cells on a Tet-ON reporter system.
Improvements on the efficiency
Whereas genome-editing by the catalytically active Cas9 nuclease can be accompanied by irreversible off-target genomic alterations, CRISPRi is highly specific with minimal off-target reversible effects for two distinct sgRNA sequences. Nonetheless, several methods have been developed to improve the efficiency of transcriptional modulation.
SunTag protein scaffold
Recruitment of multiple copies of a protein to a target substrate (e.g., DNA, RNA, or protein) presents a general principle for signal amplification in biological systems. A repeating peptide array termed SunTag has been shown to recruit multiple copies of antibody-fusion protein and up to 24 copies of GFP. SunTag provides a versatile platform for multimerizing proteins on a target protein scaffold and may have applications in single molecule imaging and controlling biological outputs.
Other methods
Along with other improvements mentioned, factors such as the distance from the transcription start and the local chromatin state may be critical parameters in determining activation/repression efficiency. Optimization of dCas9 and sgRNA expression, stability, nuclear localization, and interaction will likely allow for further improvement of CRISPRi efficiency in mammalian cells.
Gene knockdown
A significant portion of the genome (both reporter and endogenous genes) in eukaryotes has been shown to be targetable using lentiviral constructs to express dCas9 and sgRNAs, with comparable efficiency to existing techniques such as RNAi and TALE proteins. In tandem or as its own system, CRISPRi could be used to achieve the same applications as in RNAi.
For bacteria, gene knockdown by CRISPRi has been fully implemented and characterized (off-target analysis, leaky repression) for both Gram-negative E. coli and Gram-positive B. subtilis.
Allelic series
Differential gene expression can be achieved by modifying the efficiency of sgRNA base-pairing to the target loci. In theory, modulating this efficiency can be used to create an allelic series for any given gene, in essence creating a collection of hypo- and hypermorphs. These powerful collections can be used to probe any genetic investigation. For hypomorphs, this allows the incremental reduction of gene function as opposed to the binary nature of gene knockouts and the unpredictability of knockdowns. For hypermorphs, this is in contrast to the conventional method of cloning the gene of interest under promoters with variable strength.
Loci imaging
Fusing a fluorescent protein to dCas9 allows for imaging of genomic loci in living human cells. This can be used to study chromatin architecture and nuclear organization dynamics.
Stem cells
Activation of Yamanaka factors by CRISPRi has been used to induce pluripotency in human and mouse cells providing an alternative method to iPS technology. In addition, large-scale activation screens could be used to identify proteins that promote induced pluripotency or, conversely, promote differentiation to a specific cell lineage.
Genetic screening
The ability to upregulate gene expression using dCas9-SunTag with a single sgRNA also opens the door to large-scale genetic screens, such as Perturb-seq, to uncover phenotypes that result from increased or decreased gene expression, which will be especially important for understanding the effects of gene regulation in cancer. Furthermore, CRISPRi systems have been shown to be transferable via horizontal gene transfer mechanisms such as bacterial conjugation and specific repression of reporter genes in recipient cells has been demonstrated. CRISPRi could serve as a tool for genetic screening and potentially bacterial population control.
Advantages
1. CRISPRi can silence a target gene of interest up to 99.9% repression.
2. Since CRISPRi is based on Watson-Crick base-pairing of sgRNA-DNA and an NGG PAM motif, selection of targetable sites within the genome is straightforward and flexible. Carefully defined protocols have been developed.
3. Multiple sgRNAs can not only be used to control multiple different genes simultaneously (multiplexing gene targeting), but also to enhance the efficiency of regulating the same gene target.
4. While the two systems can be complementary, CRISPRi provides advantages over RNAi. As an exogenous system, CRISPRi does not compete with endogenous machinery such as microRNA expression or function. Furthermore, because CRISPRi acts at the DNA level, one can target transcripts such as noncoding RNAs, microRNAs, antisense transcripts, nuclear-localized RNAs, and polymerase III transcripts. Finally, CRISPRi possesses a much larger targetable sequence space; promoters and, in theory, introns can also be targeted.
5. In E. coli, construction of a gene knockdown strain is extremely fast and requires only one-step oligo recombineering.
Limitations
1.The requirement of a protospacer adjacent motif (PAM) sequence limits the number of potential target sequences. Cas9 and its homologs may use different PAM sequences, and therefore could theoretically be utilized to expand the number of potential target sequences.
2.Sequence specificity to target loci is only 14 nt long (12 nt of sgRNA and 2nt of the PAM), which can recur around 11 times in a human genome. Repression is inversely correlated with the distance of the target site from the transcription start site. Genome-wide computational predictions or selection of Cas9 homologs with a longer PAM may reduce nonspecific targeting.
3.Endogenous chromatin states and modifications may prevent the sequence specific binding of dCas9-sgRNA complex. The level of transcriptional repression in mammalian cells varies between genes. Much work is needed to understand the role of local DNA conformation and chromatin in relation to binding and regulatory efficiency.
4.CRISPRi can have an influence on genes, which are in close proximity to the intended target gene. This is of special importance targeting genes, which have either overlapping neighboring genes (sense-, or antisense overlapping) or which are driven by a bidirectional promoter.