
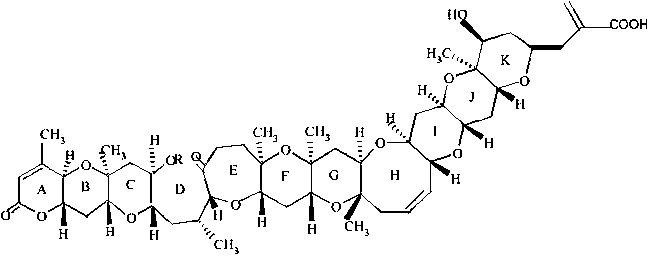
 | ||
Brevetoxin (PbTx), or brevetoxins, are a suite of cyclic polyether compounds produced naturally by a species of dinoflagellate known as Karenia brevis. Brevetoxins are neurotoxins that bind to voltage-gated sodium channels in nerve cells, leading to disruption of normal neurological processes and causing the illness clinically described as neurotoxic shellfish poisoning (NSP).
Contents
- Biosynthesis
- Impact on human and animal health
- Nitrogen and phosphorus availability vs toxicity level
- References
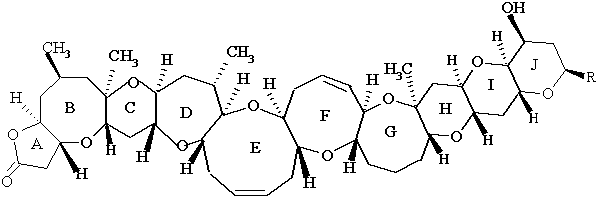
Although brevetoxins are most well-studied in K. brevis, they are also found in other species of Karenia and at least one large fish kill has been traced to brevetoxins in Chattonella.

Other Brevetoxins:

Brevetoxin-B was synthesized in 1995 by K. C. Nicolaou and coworkers in 123 steps with 91% average yield (final yield ~9·10−6) and in 2004 in a total of 90 steps with an average 93% yield for each step (0.14% overall).
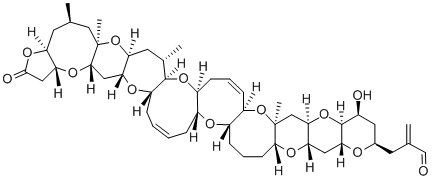
K. C. Nicolaou and coworkers reported their synthesis of Brevetoxin-1 in 1998. In 2009, Michael Crimmins and co-workers reported their synthesis of Brevetoxin-1 as well.
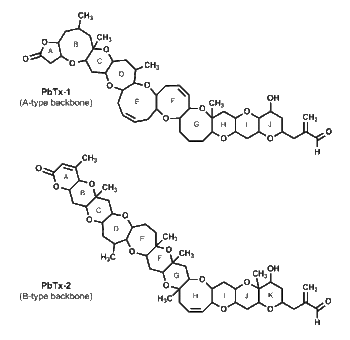
Biosynthesis
Brevetoxins share the common backbone structure of polyketides, but there are several methyl and oxygen groups that are not typical of a traditional polyketide synthesis. Labelling studies identifying the origin of the various carbon atoms have shown that the biosynthesis of brevetoxins greatly deviates from the polyketide synthetic pathway.
From the labelling experiments of Brevetoxin-B (BTX-B), a 50-carbon molecule, 16 carbon signals were enhanced by [1-C13] acetate, 30 signals were enhanced by [2-C13] acetate, and 4 carbon signals were enhanced by [methyl-C13] methionine. Furthermore, 14 intact acetate units were identified with a fifteenth two carbon unit with a weak possibility of being an acetate unit. It is clear based on the oxygen locations of BTX-B that this molecule could not be produced using a traditional polyketide synthetic pathway. Attention was turned to the citric acid cycle to solve the problem. Acetate can be used in the polyketide synthetic pathway or modified by the citric acid cycle. Intermediate products of this cycle can then be reintroduced to the polyketide synthetic pathway, resulting in the addition of atypical carbon units. Previous studies of the citric acid pathway revealed three and four carbon units that can potentially explain the atypical condensation and oxidation pattern seen in BTX-B. That being said, there is currently no explanation as to why this particular pattern is favored.
All these things considered, a proposed biosynthetic pathway for brevetoxin class compounds begins with a traditional polyketide synthesis that has the potential to incorporate larger carbon units originating from acetate modified by the citric acid cycle. After the carbon backbone is synthesized, oxidation produces the necessary epoxides that lead to the closure of the multi-ring system. It is unclear if methyl groups as seen in BTX-B are added after cyclization or during the modification of the polyketide metabolites, but it is clear that methyl groups can originate from sources outside of acetate, such as S-adenosylmethionine.
Impact on human and animal health
The potent polyether brevetoxins produced by K. brevis activate voltage-sensitive sodium channels. Specifically, brevetoxins bind to site 5 on the alpha-subunit of voltage sensitive sodium channels (VSSCs), which serve as key proteins in the structure of the cell membrane. The binding of brevetoxin to VSSCs produces three key effects: a lowering of the activation potential to more negative values, the persistent activation of the channel and therefore repetitive firing of nerves, and the inability to reverse this prolonged mean open state. This leads to a number of health problems in both humans and animals alike. The disruption of normal sodium channel function namely results in massive fish kills and the poisoning of marine mammals and other aquatic invertebrates, which in turn are a source of human health problems as well. For instance, pulmonary receptors associated with ligand-gated epithelial Na+ channels and cathepsin inhibition in macrophages have been reported to be affected by brevetoxin exposure.
The uptake of brevetoxin into both humans and animals occurs primarily through inhalation and ingestion. Dermal contact, such as through swimming in the red tides, is a suspected method of uptake, although direct contact with the toxin in the water is not well studied. In the case of inhalation, aerosolized toxins carried onshore in sea spray can cause respiratory irritation that can escalate, in more extreme cases, to more severe airway constriction, an effect observed at pM concentrations. More significant are the cases of ingestion, whether by direct swallowing of seawater during blooms of K. brevis or digestion of contaminated filter-feeding animals. After feeding upon K. brevis, aquatic invertebrates and shellfish in particular can accumulate brevetoxins, resulting in neurotoxic shellfish poisoning (NSP). In humans, the characteristic symptoms of NSP include parasthesia (tingling), reversal of hot-cold temperature sensation, myalgia (muscle pain), vertigo, ataxia (loss of coordination), abdominal pain, nausea, diarrhea, headache, bradycardia (slow heart rate), dilated pupils and as respiratory distress, as previously mentioned. The bioaccumulation effect has been observed for this toxin in the food web, and it has been noted that this accumulation is not restricted to times when K. brevis is present.
The range and degree of human health effects seems to vary annually and temporally in coastal regions, depending on the red tide density as well as variation in toxicity differences among dinoflagellate strains and their subsequent consumers. The Gulf of Mexico, and in particular the west coast of Florida, is the most heavily impacted by the adverse health and environmental effects of nearly annual K. brevis blooms. This region has suffered significant economic losses in local communities that rely on tourism and recreational fishing along with bad publicity over the years. Shellfish poisonings have been known about in Florida since the 1880s, although the cause was not identified as K. brevis until 1960.
The metabolism of brevetoxins in shellfish is particularly concerning, as certain derivatives have been shown to remain in the animal over extended periods of time. It has been shown that the main toxin produced by K. brevis, PbTx-2, is rapidly metabolized, resulting in the production of metabolites that endure in the animals system for a significantly longer period of time. This stands in contrast to PbTx-3, which is typically eliminated from the shellfish in more or less its original form within a few weeks.
Brevetoxin concentrations in seafood and the regulation of toxic substance monitoring in the animals is concerning. In Florida, only oysters and clams are monitored for NSP. Scallops are not monitored, although scallop-related NSP does not normally occur because in most cases, the muscle which does not accumulate brevetoxin to dangerous levels is consumed. Additionally, scallops are less tolerant to brevetoxins as compared to other bivalves and die off quickly after exposure to K. brevis red tides. However, smaller bivalves such as chione clams and coquinas can accumulate extremely high levels of brevetoxins and are not monitored, which could potentially impact both human and wildlife health in negative ways. According to evidence from Poli et al., whelks are implicated in an NSP event in 1996.
With respect to ichthyotoxicity, reports of massive fish kills have been reported in the Gulf of Mexico as far back as 1844. Originally, fish bioassay-guided fractionation was used to isolate the toxins, but accumulation in or food-web transfer by fish has not been regarded as a threat. Steidinger hypothesized that the presence of brevetoxin found in dolphin mortalities and prey mortalities in 1987-1988 were in part due to brevetoxin transfer through fish. While dangerous levels of brevetoxins have not been found in the muscles of live fish to date, the internal organs of fish are highly susceptible to dangerous levels of toxicity and should not be eaten. It is conjectured that chronic low-level exposure to brevetoxin metabolites can occur through shellfish and fish, although the effects of this have not been studied in detail and remain largely unknown.
Nitrogen and phosphorus availability vs toxicity level
Along the west coast of Florida, the early phase of K. brevis blooms are initiated by northerly winds, resulting in upwelling events that cause nutrients to rise towards the surface of the water and transport multiple Karenia cell species towards the shore. Here they concentrate and either continue to grow or are taken up by onshore winds that spread the cells over beaches and near shore communities. It has been shown that K. brevis blooms are limited by available nitrogen (N) or phosphorus (P), but until recently it was not clear what sources K. brevis was utilizing for these key developmental nutrients. The most likely proposition is some combination of the upwelling of subsurface nutrients, land runoff, N2-fixation, drainage from phosphate mines and atmospheric deposition provides the necessary support for the blooms.
N-limitation directly affects the growth potential of blooms and the toxicity of K. brevis cells that comprise them. When N-limitation is present, intracellular brevetoxin concentrations (fg/µm3) increased up to 2.5-fold in laboratory cultures, implying that during periods of N-limitation of algal growth, there is a higher chance of brevetoxin influx into the marine food web.
The toxin content per cell increases when algal growth becomes P-limited. Various field measurements collected in the Gulf of Mexico have shown that the brevetoxin contents of K. brevis cells cary between 1 and 68 pg/cell, however, Hardison et al. discovered that during periods of transient P- and N-limitation, there is a 2- to 5-fold increase in brevetoxins per mole cell carbon or unit of cell volume. Hardison concluded that this data suggest that the exposure of marine ecosystems to significantly different toxin levels depends on the nutrient status of the K. brevis cells. While brevetoxins remain intracellular during early stages of bloom development, the triggering of apoptosis and cell lysis with age release the toxins into the surrounding waters, implying that greater P-limitation that results in more cell death ultimately elevates brevetoxin levels. These high levels may persist in a food chain long after a bloom has subsided due to brevetoxin’s high affinity for adsorbing to biological surfaces like sea grass fronds, and thereby accumulating in consuming organisms.
Overall, brevetoxins seem to increase under N- and P-limitation, however, the concentration of brevetoxins per cell under P-limitation have been reported to be roughly twice those under N-limitation. One major concern of this is that management of shellfish bed closures operating under the assumption that brevetoxin concentrations per cell do not vary may compromise public safety if a bloom became nutrient limited.