
 | ||
Antimicrobial peptides (AMPs), also called host defense peptides (HDPs) are part of the innate immune response found among all classes of life. Fundamental differences exist between prokaryotic and eukaryotic cells that may represent targets for antimicrobial peptides. These peptides are potent, broad spectrum antibiotics which demonstrate potential as novel therapeutic agents. Antimicrobial peptides have been demonstrated to kill Gram negative and Gram positive bacteria, enveloped viruses, fungi and even transformed or cancerous cells. Unlike the majority of conventional antibiotics it appears as though antimicrobial peptides may also have the ability to enhance immunity by functioning as immunomodulators.
Contents
- Structure
- Activities
- Immunomodulation
- Mode of action
- Therapeutic potential
- Anticancer
- Selectivity
- Factors
- Mechanism
- Control
- Bacterial resistance
- Examples
- Bioinformatics
- References
Marine fish sources have high levels of antimicrobial compounds with in vivo testing confirming the efficacy of fish peptides used in food/feed ingredients.
Structure
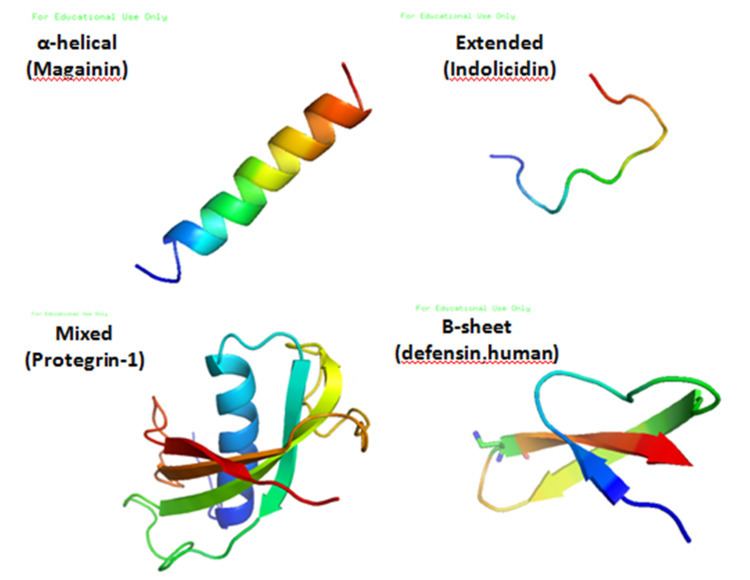
Antimicrobial peptides are a unique and diverse group of molecules, which are divided into subgroups on the basis of their amino acid composition and structure. Antimicrobial peptides are generally between 12 and 50 amino acids. These peptides include two or more positively charged residues provided by arginine, lysine or, in acidic environments, histidine, and a large proportion (generally >50%) of hydrophobic residues. The secondary structures of these molecules follow 4 themes, including i) α-helical, ii) β-stranded due to the presence of 2 or more disulfide bonds, iii) β-hairpin or loop due to the presence of a single disulfide bond and/or cyclization of the peptide chain, and iv) extended. Many of these peptides are unstructured in free solution, and fold into their final configuration upon partitioning into biological membranes. It contains hydrophilic amino acid residues aligned along one side and hydrophobic amino acid residues aligned along the opposite side of a helical molecule. This amphipathicity of the antimicrobial peptides allows them to partition into the membrane lipid bilayer. The ability to associate with membranes is a definitive feature of antimicrobial peptides although membrane permeabilization is not necessary. These peptides have a variety of antimicrobial activities ranging from membrane permeabilization to action on a range of cytoplasmic targets.
Activities
The modes of action by which antimicrobial peptides kill microbes are varied, and may differ for different bacterial species. Some antimicrobial peptides kill both bacteria and fungi, e.g., psoriasin kills E. coli and several filamentous fungi. The cytoplasmic membrane is a frequent target, but peptides may also interfere with DNA and protein synthesis, protein folding, and cell wall synthesis. The initial contact between the peptide and the target organism is electrostatic, as most bacterial surfaces are anionic, or hydrophobic, such as in the antimicrobial peptide Piscidin. Their amino acid composition, amphipathicity, cationic charge and size allow them to attach to and insert into membrane bilayers to form pores by ‘barrel-stave’, ‘carpet’ or ‘toroidal-pore’ mechanisms. Alternately, they may penetrate into the cell to bind intracellular molecules which are crucial to cell living. Intracellular binding models includes inhibition of cell wall synthesis, alteration of the cytoplasmic membrane, activation of autolysin, inhibition of DNA, RNA, and protein synthesis, and inhibition of certain enzymes. However, in many cases, the exact mechanism of killing is not known. One emerging technique for the study of such mechanisms is dual polarisation interferometry. In contrast to many conventional antibiotics these peptides appear to be bactericidal instead of bacteriostatic. In general the antimicrobial activity of these peptides is determined by measuring the minimal inhibitory concentration (MIC), which is the lowest concentration of drug that inhibits bacterial growth.
Immunomodulation
In addition to killing bacteria directly they have been demonstrated to have a number of immunomodulatory functions that may be involved in the clearance of infection, including the ability to alter host gene expression, act as chemokines and/or induce chemokine production, inhibiting lipopolysaccharide induced pro-inflammatory cytokine production, promoting wound healing, and modulating the responses of dendritic cells and cells of the adaptive immune response. Animal models indicate that host defence peptides are crucial for both prevention and clearance of infection. It appears as though many peptides initially isolated as and termed "antimicrobial peptides" have been shown to have more significant alternative functions in vivo (e.g. hepcidin).
Mode of action
Several methods have been used to determine the mechanisms of antimicrobial peptide activity. In particular, solid-state NMR studies have provided an atomic-level resolution explanation of membrane disruption by antimicrobial peptides.
Therapeutic potential
These peptides are excellent candidates for development as novel therapeutic agents and complements to conventional antibiotic therapy because they generally have a broad range of activity, are bactericidal as opposed to bacteriostatic and require a short contact time to induce killing. A number of naturally occurring peptides and their derivatives have been developed as novel anti-infective therapies for conditions as diverse as oral mucositis, lung infections associated with cystic fibrosis (CF), cancer, and skin and wound infections. Pexiganan has been shown to be useful to treat infection related diabetic foot ulcer.
A major limitation to the therapeutic potential is the possibility of bacteria developing resistance to the peptides, and particularly if that produces a resistance to the body's own immune system use of those peptides. That is, providing a lot of the peptides as a therapeutic agent makes it easier for resistance to evolve; unlike antibiotic resistance, however, resistance to antimicrobial peptides mimicking those produced by humans can make the bacteria more resistant to the body's own immune system rather than just the antibiotic.
Antimicrobial peptides have been successively incorporated into topical therapeutics. A major challenge associated with systemic delivery of an antimicrobial peptides is their susceptibility to proteolytic degradation. That is, the peptides are quickly broken down when introduced in the bloodstream.
Anticancer
Some of the cecropins (e.g. cecropin A, and cecropin B) have anticancer properties and are called anticancer peptides (ACPs). Hybrid ACPs based on Cecropin A have been studied for anticancer properties.
Selectivity
In the competition of bacterial cells and host cells with the antimicrobial peptides, antimicrobial peptides will preferentially interact with the bacterial cell to the mammalian cells, which enables them to kill microorganisms without being significantly toxic to mammalian cells. Selectivity is a very important feature of the antimicrobial peptides and it can guarantee their function as antibiotics in host defense systems.
Factors
There are some factors that are closely related to the selectivity property of antimicrobial peptides, among which the cationic property contributes most. Since the surface of the bacterial membranes is more negatively charged than mammalian cells, antimicrobial peptides will show different affinities towards the bacterial membranes and mammalian cell membranes.
In addition, there are also other factors that will affect the selectivity. It’s well known that cholesterol is normally widely distributed in the mammalian cell membranes as a membrane stabilizing agents but absent in bacterial cell membranes; and the presence of these cholesterols will also generally reduce the activities of the antimicrobial peptides, due either to stabilization of the lipid bilayer or to interactions between cholesterol and the peptide. So the cholesterol in mammalian cells will protect the cells from attack by the antimicrobial peptides.
Besides, the transmembrane potential is well-known to affect peptide-lipid interactions. There's an inside-negative transmembrane potential existing from the outer leaflet to the inner leaflet of the cell membranes and this inside-negative transmembrane potential will facilitate membrane permeabilization probably by facilitating the insertion of positively charged peptides into membranes. By comparison, the transmembrane potential of bacterial cells is more negative than that of normal mammalian cells, so bacterial membrane will be prone to be attacked by the positively charged antimicrobial peptides.
Similarly, it is also believed that increasing ionic strength, which in general reduces the activity of most antimicrobial peptides, contributes partially to the selectivity of the antimicrobial peptides by weakening the electrostatic interactions required for the initial interaction.
Mechanism
The cell membranes of bacteria are rich in acidic phospholipids, such as phosphatidylglycerol and cardiolipin. These phospholipid headgroups are heavily negatively charged. Therefore, the outmost leaflets of the bilayer which is exposed to the outside of the bacterial membranes are more attractive to the attack of the positively charged antimicrobial peptides. So the interaction between the positive charges of antimicrobial peptides and the negatively charged bacterial membranes is mainly the electrostatic interactions, which is the major driving force for cellular association. In addition, since antimicrobial peptides form structures with a positively charged face as well as a hydrophobic face, there are also some hydrophobic interactions between the hydrophobic regions of the antimicrobial peptides and the zwitterionic phospholipids (electrically neutral) surface of the bacterial membranes, which act only as a minor effect in this case.
In contrast, the outer part of the membranes of plants and mammals is mainly composed of lipids without any net charges since most of the lipids with negatively charged headgroups are principally sequestered into the inner leaflet of the plasma membranes. Thus in the case of mammalian cells, the outer surfaces of the membranes are usually made of zwitterionic phosphatidylcholine and sphingomyelin, even though a small portion of the membrane's outer surfaces contain some negatively charged gangliosides. Therefore, the hydrophobic interaction between the hydrophobic face of amphipathic antimicrobial peptides and the zwitterionic phospholipids on the cell surface of mammalian cell membranes plays a major role in the formation of peptide-cell binding. However, the hydrophobic interaction is relatively weak when compared to the electrostatic interaction, thus, the antimicrobial peptides will preferentially interact with bacterial membranes.
Dual polarisation interferometry has been used in vitro to study and quantify the association to headgroup, insertion into the bilayer, pore formation and eventual disruption of the membrane.
Control
A lot of effort has been put into controlling cell selectivity. For example, Katsumi tried to modify and optimize the physicochemical parameters of the peptides to control the selectivities, including net charge, helicity, hydrophobicity per residue (H), hydrophobic moment (μ) and the angle subtended by the positively charged polar helix face (Φ). Other mechanisms like the introduction of D-amino acids and fluorinated amino acids in the hydrophobic phase are believed to break the secondary structure and thus reduce hydrophobic interaction with mammalian cells. Wan L Z, el found that Pro→Nlys substitution in Pro-containing β-turn antimicrobial peptides was a promising strategy for the design of new small bacterial cell-selective antimicrobial peptides with intracellular mechanisms of action. Nadezhda V el suggested that direct attachment of magainin to the substrate surface decreased nonspecific cell binding and led to improved detection limit for bacterial cells such as Salmonella and E. coli.
Bacterial resistance
Bacteria use various resistance strategies to avoid antimicrobial peptide killing. Some microorganisms alter net surface charges. Staphylococcus aureus transports D-alanine from the cytoplasm to the surface teichoic acid which reduces the net negative charge by introducing basic amino groups. S. aureus also modifies its anionic membranes via MprF with L-lysine, increasing the positive net charge. The interaction of antimicrobial peptides with membrane targets can be limited by capsule polysaccharide of Klebsiella pneumoniae. Alterations occur in Lipid A. Salmonella species reduce the fluidity of their outer membrane by increasing hydrophobic interactions between an increased number of Lipid A acyl tails by adding myristate to Lipid A with 2-hydroxymyristate and forming hepta-acylated Lipid A by adding palmitate. The increased hydrophobic moment is thought to retard or abolish antimicrobial peptide insertion and pore formation. The residues undergo alteration in membrane proteins. In some Gram-negative bacteria, alteration in the production of outer membrane proteins correlates with resistance to killing by antimicrobial peptides. Nontypeable Hemophilus influenzae transports AMPs into the interior of the cell, where they are degraded. And H. influenzae remodels its membranes to make it appear as if the bacterium has already been successfully attacked by AMPs, protecting it from being attacked by more AMPs. ATP-binding cassette transporters import antimicrobial peptides and the resistance-nodulation cell-division efflux pump exports antimicrobial peptides. Both transporters have been associated with antimicrobial peptide resistance. Bacteria produce proteolytic enzymes,which may degrade antimicrobial peptides leading to their resistance. Outer membrane vesicles produced by Gram-negative bacteria bind the antimicrobial peptides and sequester them away from the cells, thereby protecting the cells. The outer membrane vesicles are also known to contain various proteases, peptidases and other lytic enzymes, which may have a role in degrading the extracellular peptide and nucleic acid molecules, which if allowed to reach to the bacterial cells may be dangerous for the cells. Cyclic-di-GMP signaling had also been involved in the regulation of antimicrobial peptide resistance in Pseudomonas aeruginosa
While these examples show that resistance can evolve naturally, there is increasing concern that using pharmaceutical copies of antimicrobial peptides can make resistance happen more often and faster. In some cases, resistance to these peptides used as a pharmaceutical to treat medical problems can lead to resistance, not only to the medical application of the peptides, but to the physiological function of those peptides. Further research is needed to determine if this will lead to greater harm than benefit from the use of certain antimicrobial peptides.
Examples
Antimicrobal peptides are produced by all known species, including peptides from bacteria, from fungi, from hydra, insects (mastoparan, poneratoxin, cecropin, moricin, melittin and others), frogs (magainin, dermaseptin and others), and mammals (for example, cathelicidins, defensins and protegrins). Research has increased in recent years to develop artificially-engineered mimics of antimicrobial peptides such as SNAPPs, in part due to the prohibitive cost of producing naturally-derived AMPs.
Bioinformatics
Several bioinformatic databases exist to catalogue antimicrobial peptides such as CAMP, CAMP release 2 (Collection of sequences and structures of antimicrobial peptides) the Antimicrobial Peptide Database (http://aps.unmc.edu/AP/main.php), LAMP, BioPD (see external links) and ADAM (A Database of Anti-Microbial peptides) (http://bioinformatics.cs.ntou.edu.tw/adam/). The Antimicrobial peptide databases may be divided into two categories on the basis of the source of peptides it contains, as specific databases and general databases. These databases have various tools for antimicrobial peptides analysis and prediction. For example, CAMP contains AMP prediction, feature calculator, BLAST search, clustalW, VAST, PRATT, Helical wheel etc. In addition, ADAM allows users tosearch or browse through AMP sequence-structure relationships.