
Rank Species | Higher classification Aliivibrio | |
 | ||
Similar | ||
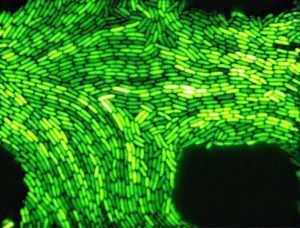
Aliivibrio fischeri is a gram-negative, rod-shaped bacterium found globally in marine environments. A. fischeri has bioluminescent properties, and is found predominantly in symbiosis with various marine animals, such as the bobtail squid. It is heterotrophic and moves by means of flagella. Free-living A. fischeri cells survive on decaying organic matter. The bacterium is a key research organism for examination of microbial bioluminescence, quorum sensing, and bacterial-animal symbiosis. It is named after Bernhard Fischer, a German microbiologist.
Contents

rRNA comparison led to the reclassification of this species from genus Vibrio to the newly created Aliivibrio in 2007.
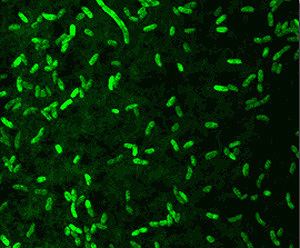
Ecology
Planktonic A. fischeri bacteria are found in very low quantities (almost undetectable) in almost all oceans of the world, preferentially in temperate and subtropical waters. These free-living bacteria subsist on organic matter in the water. They are found in higher concentrations in symbiosis with certain deep sea life within photophores (special light organs); or as part of the normal gut microbiota of marine animals.
Symbiotic relationship

A. fischeri have symbiotic relationships with several marine animals. The most prolific of these relationships is with the Hawaiian bobtail squid (Euprymna scolopes). A. fischeri inhabit photophores in the mantle of the Hawaiian bobtail squid.
A. fischeri cells in the ocean inoculate the light organs of juvenile squid and fish. Ciliated cells within the animals' photophores (light-producing organs) selectively draw in the symbiotic bacteria. These cells promote the growth of the symbionts and actively reject any competitors. The bacteria cause these cells to die off once the light organ is sufficiently colonised.
The light organs of certain squid contain reflective plates that intensify and direct the light produced, due to proteins known as reflectins. They regulate the light for counter-illumination camouflage, requiring the intensity to match that of the sea surface above. Sepiolid squid expel 90% of the symbiotic bacteria in their light organ each morning in a process known as "venting". Venting is thought to provide the free-living inoculum source for newly hatched squid.
The bioluminescence of A. fischeri is caused by transcription of the lux operon, which is induced through population-dependent quorum sensing. The population of A. fischeri needs to reach an optimal level to activate the lux operon and stimulate light production. The circadian rhythm controls light expression, where luminescence is much brighter during the day and dimmer at night, as required for camouflage.
Genetics of bioluminescence
The bacterial luciferin-luciferase system is encoded by a set of genes labelled the Lux operon. In A. fischeri, five such genes (LuxCDABEG) have been identified as active in the emission of visible light, and two genes (LuxR and LuxI) are involved in regulating the operon. Several external and intrinsic factors appear to either induce or inhibit the transcription of this gene set and produce or suppress light emission.
A. fischeri is one of many species of bacteria that commonly form symbiotic relationships with marine organisms. Marine organisms contain bacteria that use bioluminescence so they can find mates, ward off predators, attract prey, or communicate with other organisms. In return, the organism the bacteria are living within provides the bacteria with a nutrient-rich environment. The Lux operon is a 9-kilobase fragment of the A. fischeri genome that controls bioluminescence through the catalyzation of the enzyme luciferase. This operon has a known gene sequence of luxCDAB(F)E, where lux A and lux B code for the components of luciferase, and the lux CDE codes for a fatty acid reductase complex that makes the fatty acids necessary for the luciferase mechanism. Lux C codes for the enzyme acyl-reductase, lux D codes for acyl-transferase, and lux E makes the proteins needed for the enzyme acyl-protein synthetase. Luciferase produces blue/green light through the oxidation of reduced flavin mononucleotide and a long-chain aldehyde by diatomic oxygen. The reaction is summarized as: FMNH2+O2+R-CHO → FMN + R-COOH + H2O + light
The reduced flavinmononucleotide (FMNH) is provided by the fre gene, also referred to as LuxG. In A. fischeri, it is directly next to LuxE (giving LuxCDABE-fre) from 1042306 to 1048745 [1]
To generate the aldehyde needed in the reaction above, three additional enzymes are needed. The fatty acids needed for the reaction are pulled from the fatty acid biosynthesis pathway by acyl-transferase. Acyl-transferase reacts with acyl-ACP to release R-COOH, a free fatty acid. R-COOH is reduced by a two-enzyme system to an aldehyde. The reaction is: R-COOH+ATP+NADPH→ R-CHO+AMP+PP+NADP+ (Winfrey et al., 1997). Although the lux operon encodes the enzymes necessary for the bacteria to glow, bioluminescence is regulated by autoinduction. An autoinducer is a transcriptional promoter of the enzymes necessary for bioluminescence. Before the glow can be luminized, a certain concentration of an autoinducer must be present. So, for bioluminescence to occur, high colony concentrations of A. fischeri should be present in the organism.