
 | ||
The turtle shell is a highly complicated shield for the ventral and dorsal parts of turtles, tortoises and terrapins (all classified as "turtles" by zoologists), completely enclosing all the vital organs of the turtle and in some cases even the head. It is constructed of modified bony elements such as the ribs, parts of the pelvis and other bones found in most reptiles. The bone of the shell consists of both skeletal and dermal bone, showing that the complete enclosure of the shell probably evolved by including dermal armor into the rib cage.
Contents
- Shell nomenclature
- Carapace
- Plastron
- Plastral formula
- Scutes
- Septicemic cutaneous ulcerative disease SCUD
- Shell rot
- Pyramiding
- Broken shells
- References
The shell of the turtle is an important study, not just because of the obvious protection it provides for the animal, but also as an identification tool, in particular with fossils as the shell is one of the likely parts of a turtle to survive fossilization. Hence understanding the structure of the shell in living species gives us comparable material with fossils.
The shell of the hawksbill turtle, among other species, has been used as a material for a wide range of small decorative and practical items since antiquity, but is normally referred to as tortoiseshell.
Shell nomenclature
The turtle shell is made up of numerous bony elements, generally named after similar bones in other vertebrates, and a series of keratinous scutes which are also uniquely named. Some of those bones that make the top of the shell, carapace, evolved from the scapula rami of the clavicles along with the dorsal and superficial migration of the clecthra. The ventral surface is called the plastron. These are joined by an area called the bridge. The actual suture between the bridge and the plastron is called the anterior bridge strut. In Pleurodires the posterior pelvis is also part of the carapace, fully fused with it. This is not the case in Cryptodires which have a floating pelvis. The anterior bridge strut and posterior bridge strut are part of the plastron, on the carapace are the sutures into which they insert, known as the Bridge carapace suture.
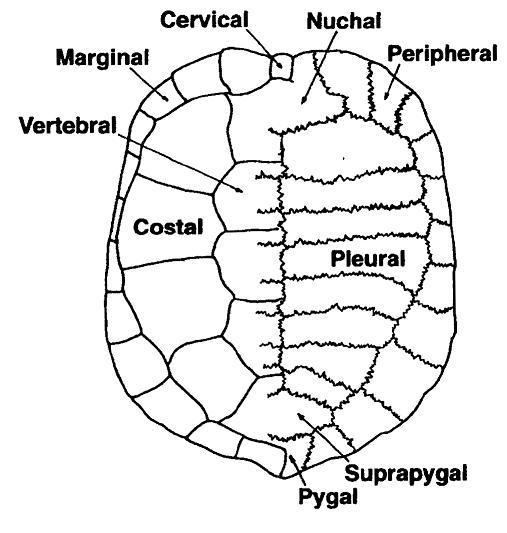
The bones of the shell are named for standard vertebrate elements. As such the carapace is made up of 8 pleurals on each side, these are a combination of the ribs and fused dermal bone. Outside of this at the anterior of the shell is the single nuchal bone, a series of 11 paired periphals then extend along each side. At the posterior of the shell is the pygal bone and in front of this nested behind the eighth pleurals is the suprapygal.
Between each of the pleurals are a series of neural bones, which although always present are not always visible, in many species of Pleurodire they are submerged below the pleurals. Beneath the neural bone is the Neural arch which forms the upper half of the encasement for the spinal chord. Below this the rest of the vertebral column. Some species of turtles have some extra bones called mesoplastra, which are located between the carapace and plastron in the bridge area. They are present in most Pelomedusid turtles.
The skeletal elements of the plastron are also largely in pairs. Anteriorly there are two epiplastra, with the hyoplastra behind them. These enclose the singuar entoplastron. These make up the front half of the plastron and the hyoplastron contains the anterior bridge strut. The posterior half is made up of two hypoplastra (containing the posterior bridge strut) and the rear is a pair of xiphiplastra.
Overlying the boney elements are a series of scutes, which are made of keratin and are a lot like horn or nail tissue. In the center of the carapace are 5 vertebral scutes and out from these are 4 pairs of costal scutes. Around the edge of the shell are 12 pairs of marginal scutes. All these scutes are aligned so that for the most part the sutures between the bones are in the middle of the scutes above. At the anterior of the shell there may be a cervical scute (sometimes incorrectly called a nuchal scute) however the presence or absence of this scute is highly variable, even within species.
On the plastron there are two gular scutes at the front, followed by a pair of pectorals, then abdominals, femorals and lastly anals. A particular variation is the Pleurodiran turtles have an intergular scute between the gulars at the front, giving them a total of 13 plastral scutes. Compared to the 12 in all Cryptodiran turtles.
Carapace
The carapace is the dorsal (back), convex part of the shell structure of a turtle, consisting of the animal's ossified ribs fused with the dermal bone. The spine and expanded ribs are fused through ossification to dermal plates beneath the skin to form a hard shell. Exterior to the skin the shell is covered by scutes, which are horny plates made of keratin that protect the shell from scrapes and bruises. A keel, a ridge that runs from front to the back of the animal is present in some species, these may be single, paired or even three rows of them. In most turtles the shell is relatively uniform in structure, species variation in general shape and color being the main differences. However the soft shell turtles, pig-nose turtles and the leatherback sea turtle have lost the scutes and reduced the ossification of the shell. This leaves the shell covered only by skin. These are all highly aquatic forms.
The evolution of the turtle's shell is unique because of how the carapace represents transformed vertebrae and ribs. While other tetrapods have their scapula, or shoulder blades, found outside of the ribcage, the scapula for turtles is found inside the ribcage. The shells of other tetrapods, such as armadillos, are not linked directly to the vertebral column or rib cage allowing the ribs to move freely with the surrounding intercostal muscle. However, analysis of the transitional fossil, Eunotosaurus africanus shows that early ancestors of turtles lost that intercostal muscle usually found between the ribs.
Recent breakthroughs in stem-turtle fossil records contribute to the study of the evolution of the turtle's shell. The first piece of fossil record discovered, essential for building the evolution and development model, was Germany and Thailand’s 214-million-year-old Late Triassic reptile Proganochelys, which marked as the first point of full shell development and carapace ossification in Testudines.
The following phenomenal discovery of China’s 220-million-year-old stem turtle, predating the Proganonchelys by 6 million years, the Odontochelys semitestacea fossil in China shed light on the intermediate stages of turtle carapace evolution by exhibiting a partially formed dorsal carapace. One major discovery was that O. semitestacea provided documentation that the plastron evolved before the carapace structure. Close examination of the partially formed carapace revealed similarity to the fully formed carapace in crown turtles such as the lack of intercostal muscles and limited rib mobility. Furthermore, it is made up of modified forms of laterally expanded and broadened ribs without ossification, similar in structure to the modern turtle embryo.
The addition of South Africa’s 260-million-year-old Permian stem reptile E. africanus, the hypothesized earliest stem turtle predating the O. semitestacea by 40 million years, revealed the early stages of carapace evolution. Histological data characterized E. africanus and Proganochelys ribs as homologous structures, both broadened and T-shaped in cross-section.
Lyson hypothesizes that this morphology in E. africanus suggests that turtles may have fossorial origin. The wide torso gave rise to the turtle shell but during the Permian period, the broadened ribs may have provided great stability in burrowing. The skeletal structure of E. africanus in comparison to the extant fossorial gopher tortoise share similar features adapted to withstand the impact and force needed in digging. For example, E. africanus exhibits shoulders and forelimb adapted to burrowing, showing increased muscle indicated in structures such as their tubercle on the posterior coracoid and their large and wide terminal phalanges. Furthermore, fossoriality may have helped E. africanus survive the global mass extinction which wiped out over ninety percent of species at the end of the Permian period.
The most recent finding of a Middle Triassic stem-turtle offers both a morphological and temporal intermediate to the E. africanus and O. semitestacea, contributing to the construction of carapace evolution timeline. The 240 million-year-old Pappochelys fossil found in Germany shows similarly broadened and T-shaped ribs that vary in shape with respect to position along the spine.
The carapacial ridge has been found to play an essential role in the development of the turtle shell. Embryological analyses show that the carapacial ridge initiates the formation of the turtle shell. It causes axial arrest which causes the ribs to be dorsalized, the shoulder girdle to be rearranged and encapsulated in the rib cage, and the carapace to develop. O. semitestacea presents evidence of axial arrest that is observed in embryos but lacks fan-shaped ribs and a carapace. This suggests that the primitive carapacial ridge functioned differently and must have gained the function of mediating the ribs and carapace development later.
Plastron
The plastron is the nearly flat part of the shell structure of a turtle, what one would call the belly or ventral surface of the shell. It also includes within its structure the anterior and posterior bridge struts and the bridge of the shell. The plastron is made up of nine bones and the two epiplastra at the anterior border of the plastron are homologous to the clavicles of other tetrapods. The rest of the plastral bones are homologous to the abdominal ribs of other tetrapods.
The evolution of the plastron has remained more mysterious, though Georges Cuvier, a French naturalist and zoologist in the 19th century, wrote that the plastron developed primarily from the sternum of the turtle. This fits well with the knowledge obtained through embryological studies, showing that changes in the pathways of rib development often result in malformation or loss of the plastron. This phenomenon occurs in turtle development, but instead of experiencing complete loss of the sternum the turtle body plan repurposes the bone into the form of the plastron.
The discovery of an ancestral turtle fossil, Pappochelys rosinae, provides additional clues as to how the plastron formed. Pappochelys serves as an intermediate form between two early stem-turtles, E. africanus and Odontochelys, the latter of which possesses a fully formed plastron. In place of a modern plastron, Pappochelys has paired gastralia, like those found in E. africanus. Pappochelys is different from its ancestor because the gastralia show signs of having once been fused, as indicated by the fossil specimens which show forked ends. This evidence shows a gradual change from paired gastralia, to paired and fused gastralia, and finally to the modern plastron across these three specimens.
In certain families there is a hinge between the pectoral and abdominal scutes allowing the turtle to almost completely enclose itself. In certain species the sex of a testudine can be told by whether the plastron is concave, male or convex, female. This is because of the mating position; the male's concave plastron allows it to more easily mount the female during copulation.
The plastral scutes join along a central seam down the middle of the plastron. The relative lengths of the seam segments can be used to help identify a species of turtle. There are six laterally symmetric pairs of scutes on the plastron: gular, humeral, pectoral, abdominal, femoral, and anal (going from the head to the tail down the seam); the abdominal and gular scute seams are approximately the same length, and the femoral and pectoral seams are approximately the same length.
The gular scute or gular projection on a turtle is the most anterior part of the plastron, the underside of the shell. Some tortoises have paired gular scutes, while others have a single undivided gular scute. The gular scutes may be referred to as a gular projection if they stick out like a trowel.
Plastral formula
The plastral formula is used to compare the sizes of the individual plastral scutes (measured along the midseam). The following plastral scutes are often distinguished (with their abbreviation):
Comparison of the plastral formulas provides distinction between the two species. For example, for the eastern box turtle, the plastral formula is: an > abd > gul > pect > hum >< fem
Turtle plastrons were used by the ancient Chinese in a type of divination called plastromancy. See also oracle bones.
Scutes
The turtle's shell is covered in scutes that are made of keratin. The individual scutes as shown above have specific names and are generally consistent across the various species of turtles. Terrestrial tortoises do not shed their scutes. New scutes grow by the addition of keratin layers to the base of each scute. Aquatic chelonii shed individual scutes. The scute effectively forms the skin over the underlying bony structures; there is a very thin layer of subcutaneous tissue between the scute and the skeleton. The scutes can be brightly colored in some species, but the basal color is a grey to dark brown color dorsally; the plastral scutes are often white to yellow in base color. A turtle's shell consists of three rows of scutes, numbering 4, 9, and 25 from the inside out. These are the squares of the first three prime numbers, viz. 22, 32, and 52. Moustakas-Verho and Cherepanov's embryological study reveals that the patterning of the plastral scutes appear independent from the patterning of carapacial scutes, suggesting that the carapace and plastron evolved separately.
Septicemic cutaneous ulcerative disease (SCUD)
Originally described by Kaplan (1957) is often known by its acronym SCUD. This disease is known to be caused by a variety of bacteria entering through some sort of abrasion, combined with poor animal husbandry. The disease is identified by its progression and what starts as ulcerative lesions of the plastron leads to a septacemic infection causing the degradation of the liver and other organs. Without treatment, this will lead to death.
Shell rot
Ulcers of the shell may be superficial or deep, and may be termed "shell rot". Ulcers are generally a result of poor husbandry. Turtles with ulcerative shell lesions should be examined and treated by a veterinarian, as the ulcers may become infected and penetrate through the shell. The shell will need to be cleaned daily, and dead tissue removed. Topical and/or injectable antibiotics are required in the case of bacterial infections. Deep ulcers may need to be repaired through surgery and the application of acrylic or fiberglass material. In a disease called "septicemic cutaneous ulcerative disease," or SCUD, ulcers may be seen on both the shell and legs. This condition is often associated with the bacteria, Citrobacter freundii.
Pyramiding
Pyramiding is a shell deformity commonly found in captive turtles, in which the shell grows unevenly resulting in a "pyramid" shape underlying each scute. This deformity can vary in severity from barely noticeable to life-threatening. Indian star tortoises and some other species display this morphology naturally.
Several causes can lead to pyramiding, including excessive animal or vegetable protein; inadequate calcium, sunlight, and/or vitamin D3; and excessive moisture / poor nutrition foods. Once pyramiding has occurred, it cannot be reversed, though if the underlying problem is corrected, the animal may still lead a long, healthy life.
Broken shells
Turtles' shells may become broken due to natural causes, accidents, or intention. When the split is not too wide the shell may be brought together by screwing bolts into the shell then connecting the bolts with a wire; otherwise, a special device may be required.