
Latin tubulus transversus | ||
 | ||
Code TH H2.00.05.2.01018TH H2.00.05.2.02013 | ||
Transverse tubules (T-tubules) are extensions of the sarcolemma (muscle cell membrane) that penetrate into the centre of skeletal and cardiac muscle cells. T-tubules are highly specialised structures, that form within the first few weeks of life, containing large amounts of specific proteins known as ion channels that allow for electrical impulses (action potentials) travelling along the sarcolemma, to enter rapidly into the cell, to initiate muscle contraction.
Contents
Structure
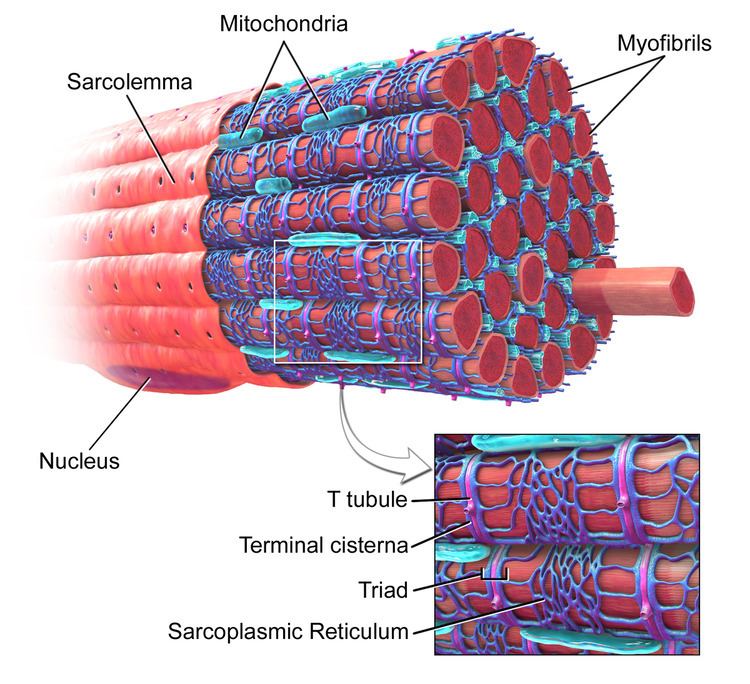
Each muscle cell is surrounded by a sarcolemma. As T-tubules are simply continuations of the sarcolemma, their membranes are very similar to that of the cell membrane, consisting of two layers of lipid (fat) molecules (known as a lipid bilayer) with proteins, including: L-type calcium channels, sodium-calcium exchangers, calcium ATPases and Beta adrenoceptors, (see below) embedded within.
Despite what their name suggests, transverse-tubules are actually networks of tubules with both transverse (running at a right angle to the sarcolemma) sections, connected by axial/longitudinal tubules. Therefore, they are also known as the transverse-axial tubular system and they run alongside myofilament bundles (proteins that are responsible for muscle contraction).
The shape of the T-tubule system is produced and maintained by a variety of proteins. For example, a protein called Amphyphysin 2 is responsible to forming the structure of the T-tubule as well as ensuring that the appropriate proteins (in particular the L-type calcium channels) are located within the T-tubule membrane. Another example, is junctophilin 2. This protein helps to form a junction between the t-tubule membrane and an intracellular calcium store known as the sarcoplasmic reticulum (SR). This junction is vital for excitation-contraction coupling within muscle cells (see below). A final example is Tcap. Tcap helps with t-tubule development. There has been shown to be increased amounts of Tcap present in response to muscles stretching. This protein is therefore responsible for an increase in the number of t-tubules as muscles grow.
Skeletal and cardiac muscles are known collectively as striated muscles. This is because of their stripey appearance due to the contractile proteins (also known as myofilaments; in paricular actin and myosin) within them.
In cardiac muscle cells, T-tubules are between 20 and 450 nanometers in diameter (nm; 1 nm=0.000000001m) and are usually located in regions called Z-discs (this is where the actin fiilaments (mentioned above) anchor, within the cell). T-tubules within the heart are closely associated with a region of the sarcoplasmic reticulum known as terminal cisternae. This is known as a dyad.
In skeletal muscle cells, T-tubules are between 20 and 40 nm in diameter and are typically located either side of the myosin strip, at the junction overlap between the A and I bands. T-tubules in skeletal muscle are associated with two terminal cisternae. This is known as a triad (see figure 1).
Excitation-contraction coupling
See Excitation-contraction coupling
T-tubules are the main sites for the coupling of excitation-contraction coupling, which is the process where an action potential, passing along the sarcolemma causes the muscle to contract. As the action potential, passes down the t-tubules, it activates the L-type calcium channels in the t-tubular membrane. In cardiac muscle activation of the L-type calcium channel, results in it forming a pore. This pore allows calcium (Ca2+) to pass into the cell, from outside. This calcium binds to and activates a receptor, known as a ryanodine receptor, which is located on the sarcoplasmic reticulum. The sarcoplasmic reticulum is a calcium store, therefore activation of the ryanodine receptor, and the subsequent opening of the RyR channel, results even more calcium flooding into the cell. This calcium is then used to produce contraction of the muscle.
In skeletal muscle cells, however, the L-type calcium channel is attached To the ryanodine receptor on the SR. Therefore, activation of the L-type calcium channel, directly activates the RyR (without the need for an influx of calcium).
Dyads and triads are, therefore, vital for excitation-contraction coupling as they allow for the L-type calcium channels and ryanodine receptors to be in close proximity, with roughly 12 nm between them.
As the t-tubules are the primary location for excitation-contraction coupling, the ion channels and proteins involved in this process are mainly situated here. For example, there are 3 times as many L-type calcium channels located within the t-tubule membrane, in comparison to the rest of the sarcolemma. Not only that but beta adrenoceptors are also highly concentrated here. Beta adrenoceptors, are receptors that are activated by adrenaline. Adrenaline is a hormone released from the adrenal gland, as part of the bodies fight or flight response. When adrenaline binds to the beta adrenoceptor, it activates it. This activation, simulates a protein called a Gs-protein, which initiates a series of reactions (known as the cyclic AMP pathway), leading to the production of Protein Kinase A (PKA). Protein Kinase A has the ability to add a phosphate to its target. In this instance one of PKAs targets is the RyR. The RyR is bound to a protein called FKBP (FK-506 binding protein), which prevents the RyR from opening. However, when the RyR becomes phosphorylated, by PKA, the FKBP unbinds, meaning that the RyR is more sensitive to stimulation. This means that there is an increased release of calcium from the SR.
As T-tubules and the sarcomplasmic reticulum spread throughout the muscle cells, they allow for synchronised calcium release across the whole cell, as the action potential is travelling so fast, that it activates all of the L-type calcium channels at almost exactly the same time. Therefore, in cells lacking t-tubules (including smooth muscle cells), the calcium that enters at the sarcolemma, has to diffuse gradually throughout the cell, activating the ryanodine receptors as a wave of calcium. This process is a much slower and results in reduced force of contraction.
Calcium control
As the t-tubular space is continuous with the extracellular space, ion concentrations between the two are very similar. However, due to the importance of the ions within the t-tubules (particularly calcium in cardiac muscle), it is very important that these concentrations remain relatively constant. As the t-tubules are very thin, they essentially trap the ions, this is important as it means that regardless of the ion concentrations elsewhere in the cell, the t-tubules still have enough ions to allow for muscle contraction. So, for example, if extracellular calcium concentration was to fall (this is known as hypocalcaemia), the t-tubule would hold on to the calcium it has. This means that cardiac excitation-contraction coupling can still occur and the heart will continue to beat.
As well as t-tubules being a site for calcium entry into the cell, they are also a site for calcium removal from the cell. This is important as it means that calcium levels within the cell can be tightly controlled in a small area (i.e. between the t-tubule and sarcoplasmic reticulum; this is known as local control). Proteins such as the Sodium(Na+)-Calcium(Ca2+) exchanger, and the sarcolemmal ATPase are located mainly in the t-tubule membrane. Both of these proteins force calcium from areas of lower concentration (i.e. within the cell) to areas of higher concentration (i.e. outside the cell; the difference in concentrations is known as the concentration gradient). This process therefore requires energy, which the proteins get from a molecule called adenosine triphosphate (ATP). The Na+-Ca2+ exchanger works by removing 1 Ca2+ from the cell in exchange for 3Na+, whereas the Ca2+ATPase works by simply removing calcium from the cell.
Remodelling
The structure of a t-tubule is vital to its function, allowing it to travel deep within the muscle cell and being in close proximity to the sarcoplasmic reticulum. Sometimes, the shape of the T-tubule is lost (it is remodelled).This occurs in the hearts of patients with heart failure. Structural changes in t-tubules can lead to the L-type calcium channels moving away from the ryanodine receptors, so that they are not in line. This can increase the time taken for calcium levels within the cell to rise and lead to weaker contractions and arrhythmia (irregular heart beat).
It has been shown that the structure of remodelled tubules can be reversed, through the use of interval training (alternating between high and low intensity workouts).
Detubulation
Scientists can uncouple T-tubules from the surface membrane using a technique known as detubulation. This relies on chemicals, such as glycerol (for skeletal muscle) or formamide (mainly for cardiac muscle), which are used due to the fact that they can't freely move across the membrane (i.e. they are osmotically active). Addition of these chemicals to the solution surrounding muscle cells causes the cells to shrink. When the chemical is withdrawn, the cells rapidly expand before returning to their normal size. It is the rapid expansion that causes the t-tubules to detach from the surface membrane. This technique can be used to investigate the function of the t-tubules.