
 | ||
RNA editing is a molecular process through which some cells can make discrete changes to specific nucleotide sequences within a RNA molecule after it has been generated by RNA polymerase. RNA editing is relatively rare, and common forms of RNA processing (e.g. splicing, 5'-capping and 3'-polyadenylation) are not usually included as editing. Editing events may include the insertion, deletion, and base substitution of nucleotides within the edited RNA molecule.
Contents
- Editing by insertion or deletion
- C to U editing
- A to I editing
- Alternative mRNA editing
- RNA editing in plant mitochondria and plastids
- RNA editing in viruses
- Origin and evolution of RNA editing
- RNA editing may be involved in RNA degradation
- References
RNA editing has been observed in some tRNA, rRNA, mRNA or miRNA molecules of eukaryotes and their viruses, archaea and prokaryotes. RNA editing occurs in the cell nucleus and cytosol, as well as within mitochondria and plastids. In vertebrates, editing is rare and usually consists of a small number of changes to the sequence of affected molecules. In other organisms, extensive editing (pan-editing) can occur; in some cases the majority of nucleotides in a mRNA sequence may result from editing.
RNA-editing processes show great molecular diversity, and some appear to be evolutionarily recent acquisitions that arose independently. The diversity of RNA editing phenomena includes nucleobase modifications such as cytidine (C) to uridine (U) and adenosine (A) to inosine (I) deaminations, as well as non-templated nucleotide additions and insertions. RNA editing in mRNAs effectively alters the amino acid sequence of the encoded protein so that it differs from that predicted by the genomic DNA sequence.
Editing by insertion or deletion
RNA editing through the addition and deletion of uracil has been found in kinetoplasts from the mitochondria of Trypanosoma brucei Because this may involve a large fraction of the sites in a gene, it is sometimes called "pan-editing" to distinguish it from topical editing of one or a few sites.
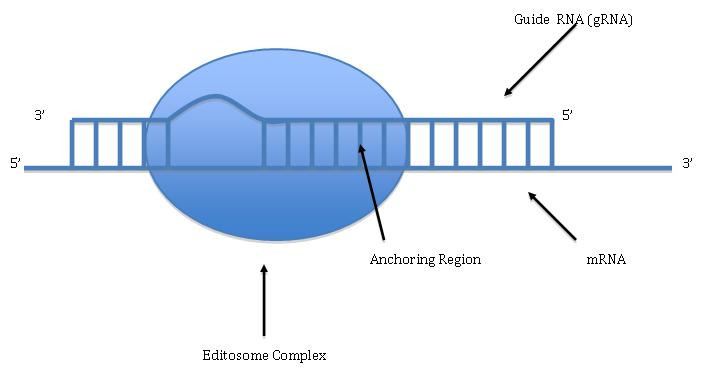
Pan-editing starts with the base-pairing of the unedited primary transcript with a guide RNA (gRNA), which contains complementary sequences to the regions around the insertion/deletion points. The newly formed double-stranded region is then enveloped by an editosome, a large multi-protein complex that catalyzes the editing. The editosome opens the transcript at the first mismatched nucleotide and starts inserting uridines. The inserted uridines will base-pair with the guide RNA, and insertion will continue as long as A or G is present in the guide RNA and will stop when a C or U is encountered. The inserted nucleotides cause a frameshift and result in a translated protein that differs from its gene.
The mechanism of the editosome involves an endonucleolytic cut at the mismatch point between the guide RNA and the unedited transcript. The next step is catalyzed by one of the enzymes in the complex, a terminal U-transferase, which adds Us from UTP at the 3’ end of the mRNA. The opened ends are held in place by other proteins in the complex. Another enzyme, a U-specific exoribonuclease, removes the unpaired Us. After editing has made mRNA complementary to gRNA, an RNA ligase rejoins the ends of the edited mRNA transcript. As a consequence, the editosome can edit only in a 3’ to 5’ direction along the primary RNA transcript. The complex can act on only a single guide RNA at a time. Therefore, a RNA transcript requiring extensive editing will need more than one guide RNA and editosome complex.
C-to-U editing
The editing involves cytidine deaminase that deaminates a cytidine base into a uridine base. An example of C-to-U editing is with the apolipoprotein B gene in humans. Apo B100 is expressed in the liver and apo B48 is expressed in the intestines. In the intestines, the mRNA has a CAA sequence edited to be UAA, a stop codon, thus producing the shorter B48 form.
C-to-U editing often occurs in the mitochondrial RNA of flowering plants. Different plants have different degrees of C-to-U editing; eight editing events occur in mitochondria of the moss Funaria hygrometrica , where as over 1700 editing events occur in the lycophytes Isoetes engelmanii. C-to-U editing is performed by members of the pentatricopeptide repeat (PPR) protein family. Angiosperms have large PPR families, acting as trans -factors for cis -elements lacking a consensus sequence; Arabidopsis has around 450 members in its PPR family. There have been a number of discoveries of PPR proteins in both plastids and mitochondria.
A-to-I editing
A-to-I editing is the main form of RNA editing in mammals and occurs in regions of double-stranded RNA (dsRNA). Adenosine deaminases acting on RNA (ADARs) are the RNA-editing enzymes involved in the hydrolytic deamination of Adenosine to Inosine (A-to-I editing). A-to-I editing can be specific (a single adenosine is edited within the stretch of dsRNA) or promiscuous (up to 50% of the adenosines are edited). Specific editing occurs within short duplexes (e.g., those formed in an mRNA where intronic sequence base pairs with a complementary exonic sequence), while promiscuous editing occurs within longer regions of duplex (e.g., pre- or pri-miRNAs, duplexes arising from transgene or viral expression, duplexes arising from paired repetitive elements). There are many effects of A-to-I editing, arising from the fact that I behaves as if it is G both in translation and when forming secondary structures. These effects include alteration of coding capacity, altered miRNA or siRNA target populations, heterochromatin formation, nuclear sequestration, cytoplasmic sequestration, endonucleolytic cleavage by Tudor-SN, inhibition of miRNA and siRNA processing, and altered splicing.
Alternative mRNA editing
Alternative U-to-C mRNA editing was first reported in WT1 (Wilms Tumor-1) transcripts, and non-classic G-A mRNA changes were first observed in HNRNPK (heterogeneous nuclear ribonucleoprotein K) transcripts in both malignant and normal colorectal samples. The latter changes were also later seen alongside non-classic U-to-C alterations in brain cell TPH2 (tryptophan hydroxylase 2) transcripts. Although the reverse amination might be the simplest explanation for U-to-C changes, transamination and transglycosylation mechanisms have been proposed for plant U-to-C editing events in mitochondrial transcripts. A recent study reported novel G-to-A mRNA changes in WT1 transcripts at two hotspots, proposing the APOBEC3A (apolipoprotein B mRNA editing enzyme, catalytic polypeptide 3A) as the enzyme implicated in this class of alternative mRNA editing. It was also shown that alternative mRNA changes were associated with canonical WT1 splicing variants, indicating their functional significance.
RNA editing in plant mitochondria and plastids
It has been shown in previous studies that the only types of RNA editing seen in the plants’ mitochondria and plastids are conversion of C-to-U and U-to-C (very rare). RNA-editing sites are found mainly in the coding regions of mRNA, introns, and other non-translated regions. In fact, RNA editing can restore the functionality of tRNA molecules. The editing sites are found primarily upstream of mitochondrial or plastid RNAs. While the specific positions for C to U RNA editing events have been fairly well studied in both the mitochondrion and plastid, the identity and organization of all proteins comprising the editosome have yet to be established. Members of the expansive PPR protein family have been shown to function as trans-acting factors for RNA sequence recognition. Specific members of the MORF (Multiple Organellar RNA editing Factor) family are also required for proper editing at several sites. As some of these MORF proteins have been shown to interact with members of the PPR family, it is possible MORF proteins are components of the editosome complex. An enzyme responsible for the trans- or deamination of the RNA transcript remains elusive, though it has been proposed that the PPR proteins may serve this function as well.
RNA editing is essential for the normal functioning of the plant’s translation and respiration activity. Editing can restore the essential base-pairing sequences of tRNAs, restoring functionality. It has also been linked to the production of RNA-edited proteins that are incorporated into the polypeptide complexes of the respiration pathway. Therefore, it is highly probable that polypeptides synthesized from unedited RNAs would not function properly and hinder the activity of both mitochondria and plastids.
C-to-U RNA editing can create start and stop codons, but it cannot destroy existing start and stop codons. A cryptic start codon is created when the codon ACG is edited to AUG.
RNA editing in viruses
RNA editing in viruses (i.e., measles, mumps, or parainfluenza) are used for stability and generation of protein variants. Viral RNAs are transcribed by a virus-encoded RNA-dependent RNA polymerase, which is prone to pausing and “stuttering” at certain nucleotide combinations. In addition, up to several hundred non-templated A's are added by the polymerase at the 3’ end of nascent mRNA. These As help stabilize the mRNA. Furthermore, the pausing and stuttering of the RNA polymerase allows the incorporation of one or two Gs or As upstream of the translational codon. The addition of the non-templated nucleotides shifts the reading frame, which generates a different protein.
Origin and evolution of RNA editing
The RNA-editing system seen in the animal may have evolved from mononucleotide deaminases, which have led to larger gene families that include the apobec-1 and adar genes. These genes share close identity with the bacterial deaminases involved in nucleotide metabolism. The adenosine deaminase of E. coli cannot deaminate a nucleoside in the RNA; the enzyme’s reaction pocket is too small to for the RNA strand to bind to. However, this active site is widened by amino acid changes in the corresponding human analog genes, APOBEC1 and ADAR, allowing deamination. The gRNA-mediated pan-editing in trypanosome mitochondria, involving templated insertion of U residues, is an entirely different biochemical reaction. The enzymes involved have been shown in other studies to be recruited and adapted from different sources. But, the specificity of nucleotide insertion via the interaction between the gRNA and mRNA are similar to the tRNA editing processes in the animal and Acanthamoeba mithochondria. Eukaryotic ribose methylation of rRNAs by guide RNA molecules is a similar form of modification.
Thus, RNA editing evolved more than once. Several adaptive rationales for editing have been suggested. Editing is often described as a mechanism of correction or repair to compensate for defects in gene sequences. However, in the case of gRNA-mediated editing, this explanation does not seem possible because if a defect happens first, there is no way to generate an error-free gRNA-encoding region, which presumably arises by duplication of the original gene region. This thinking leads to an evolutionary proposal called "constructive neutral evolution" in which the order of steps is reversed, with the gratuitous capacity for editing preceding the "defect". 31
RNA editing may be involved in RNA degradation
A study looked at the involvement of RNA editing in RNA degradation. The researchers specifically looked at the interaction between ADAR and UPF1, an enzyme involved in the nonsense-mediated mRNA decay pathway(NMD). They found that ADAR and UPF1 are found within the suprasliceosome and they form a complex that leads to the down-regulation of specific genes. The exact mechanism or the exact pathways that these two are involved in are unknown at this time. The only fact that this research has shown is that they form a complex and down-regulate specific genes.