
 | ||
The nearly neutral theory of molecular evolution is a modification of the neutral theory of molecular evolution that accounts for the fact that not all mutations are either so deleterious such that they can be ignored, or else neutral. Slightly deleterious mutations are reliably purged only when their selection coefficient is greater than one divided by the effective population size. In larger populations, a higher proportion of mutations exceed this threshold for which genetic drift cannot overpower selection, leading to fewer fixation events and so slower molecular evolution.
Contents
The nearly neutral theory was proposed by Tomoko Ohta in 1973. The population-size-dependent threshold for purging mutations has been called the "drift barrier" by Michael Lynch, and used to explain differences in genomic architecture among species.
Origins of the nearly neutral theory
According to the neutral theory of molecular evolution, the rate at which molecular changes accumulate between species should be equal to the rate of neutral mutations and hence relatively constant across species. However, this is a per-generation rate. Since larger organisms have longer generation times, the neutral theory predicts that their rate of molecular evolution should be slower. However, molecular evolutionists found that rates of protein evolution were fairly independent of generation time.
Noting that population size is generally inversely proportional to generation time, Tomoko Ohta proposed that if most amino acid substitutions are slightly deleterious, this would increase the rate of effectively neutral mutation rate in small populations, which could offset the effect of long generation times. However, because noncoding DNA substitutions tend to be more neutral, independent of population size, their rate of evolution is correctly predicted to depend on population size / generation time, unlike the rate of non-synonymous changes.
In this case, the faster rate of neutral evolution in proteins expected in small populations (due to a more lenient threshold for purging deleterious mutations) is offset by longer generation times (and vice versa), but in large populations with short generation times, noncoding DNA evolves faster while protein evolution is retarded by selection (which is more significant than drift for large populations) In 1973, Ohta published a short letter in Nature suggesting that a wide variety of molecular evidence supported the theory that most mutation events at the molecular level are slightly deleterious rather than strictly neutral.
Between then and the early 1990s, many studies of molecular evolution used a "shift model" in which the negative effect on the fitness of a population due to deleterious mutations shifts back to an original value when a mutation reaches fixation. In the early 1990s, Ohta developed a "fixed model" that included both beneficial and deleterious mutations, so that no artificial "shift" of overall population fitness was necessary. According to Ohta, however, the nearly neutral theory largely fell out of favor in the late 1980s, because the mathematically simpler neutral theory for the widespread molecular systematics research that flourished after the advent of rapid DNA sequencing. As more detailed systematics studies started to compare the evolution of genome regions subject to strong selection versus weaker selection in the 1990s, the nearly neutral theory and the interaction between selection and drift have once again become an important focus of research.
Theory
The rate of substitution,
where
where
The effect of nearly neutral mutations can depend on fluctuations in
The “fixed model” provides a slightly different explanation for the rate of protein evolution. In large
The “fixed model” expands the nearly neutral theory. Tachida classified evolution under the “fixed model” based on the product of
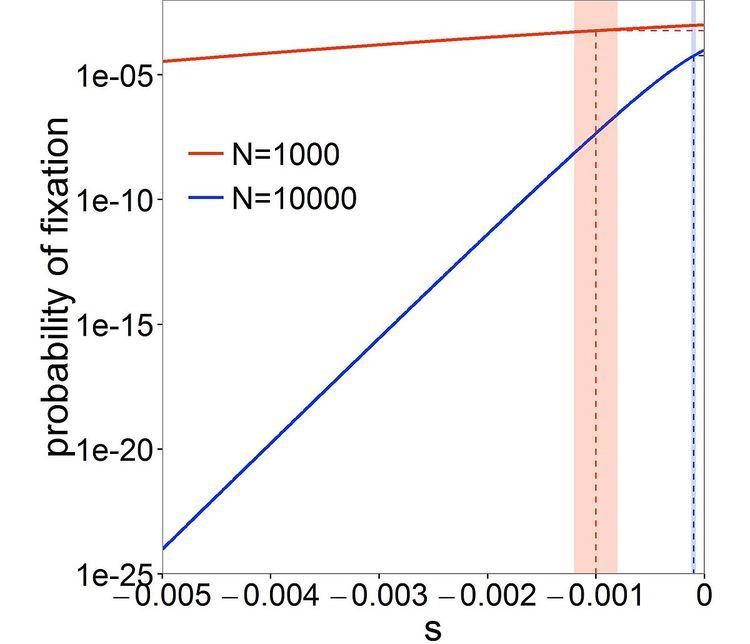
The "drift barrier" theory
Michael Lynch has proposed that variation in the ability to purge slightly deleterious mutations (i.e. variation in