
Entrez 4157 | Ensembl ENSG00000258839 | |
 | ||
External IDs | ||
Gene music using protein sequence of mc1r melanocortin 1 receptor alpha melanocyte stimulating ho
The melanocortin 1 receptor (MC1R), also known as melanocyte-stimulating hormone receptor (MSHR), melanin-activating peptide receptor, or melanotropin receptor, is a G protein–coupled receptor that binds to a class of pituitary peptide hormones known as the melanocortins, which include adrenocorticotropic hormone (ACTH) and the different forms of melanocyte-stimulating hormone (MSH).
Contents
- Gene music using protein sequence of mc1r melanocortin 1 receptor alpha melanocyte stimulating ho
- Pronounce medical words melanocortin 1 receptor
- Coloration in mammals
- Pain in mammals
- Some roles in development
- MC1R and infectioninflammation
- Role in cancer independent of skin color
- Role in kidney pathology
- Agonists
- Antagonists
- In other organisms
- Pigmentation genetics
- References
MC1R is one of the key proteins involved in regulating mammalian skin and hair color. It is located on the plasma membrane of specialized cells known as melanocytes, which produce the pigment melanin through the process of melanogenesis. It works by controlling the type of melanin being produced, and its activation causes the melanocyte to switch from generating the yellow or red phaeomelanin by default to the brown or black eumelanin in replacement.
MCR1 has also been reported to be involved in cancer (independent of skin coloration), developmental processes and susceptibilty to infections and pain.
Pronounce medical words melanocortin 1 receptor
Coloration in mammals
The MC1R protein lies within the cell membrane, and is signalled by melanocyte-stimulating hormone (MSH) released by the pituitary gland. When activated by one of the variants of MSH, typically α-MSH, MC1R initiates a complex signaling cascade that leads to the production of the brown or black pigment eumelanin. In contrast, the receptor can also be antagonized by agouti signalling peptide (ASIP), which reverts the cell back to producing the yellow or red phaeomelanin.
The pulsatile nature of ASIP signalling through MC1R produces the characteristic yellow and black agouti banding pattern observed on most mammalian hair. In some species, ASIP signaling is not of a pulsative nature, but is limited to certain regions. This is especially conspicuous in horses, where a bay horse has black legs, mane, and tail, but a reddish body. A notable exception to this is human hair, which is neither banded nor particoloured, so is thought to be regulated by α-MSH signaling through MC1R exclusively.
In the United States, about 25% of the population carries the mutated melanocortin 1 receptor that causes red hair. With one in four people as carriers, the chance of two people having a child with red hair is about 2% (one in 64). The prevalence of red hair varies considerably worldwide. People with freckles and no red hair have an 85% chance of carrying the MC1R gene that is connected to red hair. People with no freckles and no red hair have an 18% chance of carrying the MC1R gene linked to red hair.
Pain in mammals
In mutant yellow-orange mice and human redheads, both with nonfunctional MC1R, both genotypes display reduced sensitivity to noxious stimuli and increased analgesic responsiveness to morphine-metabolite analgesics. These observations suggest a role for mammalian MC1R outside the pigment cell, though the exact mechanism through which the protein can modulate pain sensation is not known.
In a certain genetic background in mice it has been reported that animals lacking MC1R had increased tolerance to capsaicin acting trough the TRPV1 receptor and decreased response to chemically induced inflammatory pain.
Humans with MC1R mutations have been reported to need approximately 20% more Inhalational anaesthetic than controls. Lidocaine was reported to be much less effective in reducing pain in another study of humans with MCR1 mutations
Some roles in development
Since G protein–coupled receptors are known to activate Signal transduction in cells, it should not be surprising to find MC1R involved in development. As one example at the cellular level, preventing signaliing by MC1R stopped erythropoiesis from proceeding from the polychromatic cell stage (poly-E in the figure) to the orthochromatic cell stage (ortho-E in the diagram). The same report showed that neutralizing antibodies to MCR1 prevented phosphorylation of STAT5 by erythropoietin, and that MCR2 and MCR5 were also involved, as shown in their model.
One example at the tissue level showed the involvement of MC1R in the normal and pathological development of articular cartilage in the mouse knee. In this study the authors compared normal mice with mice completely lacking MC1R. Even without experimental induction of osteoarthritis, mice without MC1R had less articular cartilage (as shown by the red staining in the image). After experimental induction of osteoarthritis, the defect caused by MC1R was more pronounced.
MC1R and infection/inflammation
The involvement of MC1R in a rat model of Candida albicans vaginitis was investigated. These authors suggest that MC1R is important in anti-fungal and anti-inflammatory processes, in part because siRNA knockdown of MC1R almost completely prevented the responses.
Nosocomial infections are of variable importance. One of the most important is complicated sepsis, which was defined as sepsis with organ dysfunction. One variant of MC1R (MC1RR163Q, rs885479) was reported to be associated with lowered risk of developing complicated sepsis during hospitalization after trauma. Thus, if the association is confirmed, MC1R targeting may become a therapeutic option to prevent severe sepsis.
Role in cancer independent of skin color
MC1R signalling stimulates antioxidant and DNA repair pathways, as reviewed. There are single nucleotide polymorphisms in MC1R that are associated with predisposition to nonmelanoma skin cancer. It has been reported that variants of MC1R, even in heterozygotes and independent of their effects on pigmentation, are risk factors for basal cell carcinoma and squamous cell carcinoma A review has discussed the role of some MC1R variants in melanoma and basal and squamous cell carcinomas independent of pigment production.
Role in kidney pathology
Membranous glomerulonephritis is a serious human disease that can be teated with ACTH, which is a known agonist of MC1R. In a rat model of nephritis it was found that treatment with a different agonist of MC1R improved aspects of kidney morphology and reduced proteinuria, which may help explain the benefit of ACTH in humans.
Agonists
Antagonists
In other organisms
MC1R has a slightly different function in cold-blooded animals such as fish, amphibians, and reptiles. Here, α-MSH activation of MC1R results in the dispersion of eumelanin-filled melanosomes throughout the interior of pigment cells (called melanophores). This gives the skin of the animal a darker hue and often occurs in response to changes in mood or environment. Such a physiological color change implicates MC1R as a key mediator of adaptive cryptic coloration. The role of ASIP's binding to MC1R in regulating this adaptation is unclear; however, in teleost fish at least, functional antagonism is provided by melanin-concentrating hormone. This signals through its receptor to aggregate the melanosomes toward a small area in the centre of the melanophore, resulting in the animal's having a lighter overall appearance. Cephalopods generate a similar, albeit more dramatic, pigmentary effect using muscles to rapidly stretch and relax their pigmented chromatophores. MC1R does not appear to play a role in the rapid and spectacular colour changes observed in these invertebrates.
Pigmentation genetics
MC1R gene expression is regulated by the microphthalmia-associated transcription factor (MITF). Mutations of the MC1R gene either can create a receptor that constantly signals, even when not stimulated, or can lower the receptor's activity. Alleles for constitutively active MC1R are inherited dominantly and result in a black coat colour, whereas alleles for dysfunctional MC1R are recessive and result in a light coat colour. Variants of MC1R associated with black, red/yellow, and white/cream coat colors in numerous animal species have been reported, including:
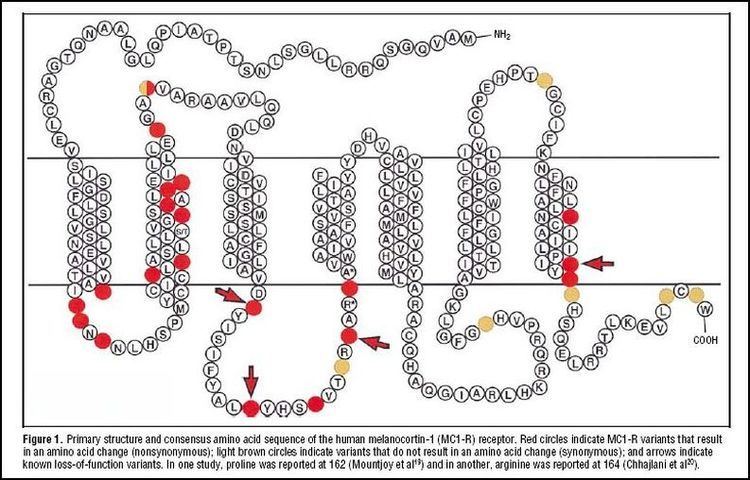
A study on unrelated British and Irish individuals demonstrated that over 80% of people with red hair and/or fair skin that tan poorly have a dysfunctional variant of the MC1R gene. This is compared to less than 20% in people with brown or black hair, and less than 4% in people showing a good tanning response.
The Out-of-Africa model proposes that modern humans originated in Africa and migrated north to populate Europe and Asia. These migrants most likely had a functional MC1R variant and, accordingly, dark hair and skin as displayed by indigenous Africans today. As humans migrated north, the absence of high levels of solar radiation in northern Europe and Asia relaxed the selective pressure on active MC1R, allowing the gene to mutate into dysfunctional variants without reproductive penalty, then propagate by genetic drift. Studies show the MC1R Arg163Gln allele has a high frequency in East Asia and may be part of the evolution of light skin in East Asian populations. No evidence is known for positive selection of MC1R alleles in Europe and there is no evidence of an association between MC1R and the evolution of light skin in European populations. The lightening of skin color in Europeans and East Asians is an example of convergent evolution.