
Domain Eukaryota Order Achromatorida Rank Genus | Subclass Haemosporidiasina Suborder Theileriina | |
 | ||
Similar Haemoproteus, Haemosporida, Hepatozoon, Haemoproteidae, Plasmodiidae | ||
2013 3 9 leucocytozoon part 1
Leucocytozoon (or Leukocytozoon) is a genus of parasitic alveolates belonging to the phylum Apicomplexa. The parasites were first seen by Vasily Danilewsky in 1884 in blood from an owl. The genus was created by Ziemann in 1898. This taxon was revised by Berestneff in 1904 and then by Sambon in 1908.
Contents
- 2013 3 9 leucocytozoon part 1
- Evolution
- Taxonomy
- Life cycle
- Diagnostic criteria
- Pathology
- Epidemiology
- Host range
- Vectors
- References
The species of this genus use blackflies (Simulium species) as their definitive host and birds as their intermediate host. There are over 100 species in this genus. Over 100 species of birds have been recorded as hosts to these parasites.
Evolution
It has been suggested that this genus arose in the late Oligocene or early Eocene at about the same time as Piciformes and Coraciiformes.
Taxonomy
The genus is divided into two subgenera: Akiba and Leucocytozoon - based on the vector species.
The only known member of the subgenus Akiba is Leucocytozoon (Akiba) caulleryi which uses members of the genus Culicoides as its vectors.
The remaining species in the genus use members of the Simulium genus as their vectors.
In 1977, Greiner and Kocan in an extensive examination of species in the order Falconiformes declared that the only valid species infecting this order was L. toddi.
L. dubreuili is considered to be restricted to the family Turdidae; L. fringillinarum is considered to be restricted to several passiform families; and L. majoris is considered restricted to the family Paridae.
L. ziemanni infects owls.
Life cycle
The life cycle of these parasites was determined in the 1930s by O'Roke in waterfowl and by Skidmore in turkeys.
Sporozoites are found in the salivary glands of an infected Simulium fly and are transmitted to the bird when the fly bites the bird to feed on its blood. The injected sporozoites find their way to the liver where they infect hepatocytes and develop into trophozoites and then into schizonts. These schizonts divide into merozoites after 4–6 days. The duration of this stage depends in part upon the species. In some species this stage may occur in the endothelial cells instead of the liver.
The nuclei of the schizonts is enclosed in a trilaminar membrane with peripherally arranged chromatin. The schizonts also contain numerous cytomers also with trilaminar membranes and containing multiple ribosomes. Repeated invagination of the cytomeres gives rise to the merozoites which also have a trilaminar membrane.
The newly released merozoites infect either erythrocytes, leukocytes, macrophages or endothelial cells. Those infecting the macrophages or endothelial cells develop into megaloschizonts. The megaloschizonts divide into primary cytomeres, which in turn multiply into smaller cytomeres, which mature into schizonts, which in turn divide into merozoites.
The merozoites have rhoptries, micronemes and three apical rings. The mitochondrion contains vesicular cristae. There may be one or two paranuclear bodies in some species. Their function is unknown.
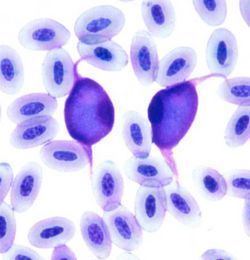
In erythrocytes or leukocytes, merozoites develop into gametocytes. Two gametocyte forms are recognised: an elongated (sail-like) form and a compact spherical form. These are usually 12-14 micrometres long. The large gametocytes tend to grossly distort the infected cells and make cell identification difficult. A pseudopigment known as volutin may be present.
The gametocytes mature in the insect midgut into macrogametocytes (female) with red-staining nuclei and microgametocytes (male) with pale-staining diffuse nuclei: these fuse to form an ookinete. The male gametocytes normally give rise to eight microgametes.
The ookinete penetrates an intestinal cell of the fly and matures into an oocyst. After several days the oocyst produces ~100 sporozoites that leave and migrate to the salivary glands of the fly.
Diagnostic criteria
Form gamonts in white blood cells and/or erythrocytes. Gametocytes cause marked enlargement and distortion of the infected cell producing a football-like appearance.
No merogony occurs in either leucocytes or erythrocytes.
Merogony occurs in the parenchyma of liver, heart, kidney, or other organs. Meronts may form large bodies divided into cytomeres.
Hemozoin deposits (pigment) are not formed - a useful distinguishing feature for Leucocytozoon from Haemoproteus and Plasmodium.
Oocysts develop rapidly in 3 – 5 days. The oocysts are small and nonexpanding, reaching 13 micrometres in diameter and typically have less than 100 short, thick sporozoites.
The vectors are Simulium or Culicoides species.
The vertebrate hosts are birds.
Type species: Leucocytozoon ziemanni
Pathology
The typical pathology of infection with these parasites includes anaemia and enlargement of the liver and spleen. Gross lesions also include pulmonary congestion and pericardial effusion.
Megaloschizonts appear as grey-white nodules found in the heart, liver, lung or spleen. Microscopically there is ischemic necrosis and associated inflammation in the heart, brain, spleen and liver due to occlusion of blood vessels by megaloschizonts in endothelial cells. Ruptured schizonts may induce granulomatous reactions in the surrounding tissues.
Clinically the majority of birds affected with leucocytozoonosis exhibit no signs. Among those that do the signs include mild to severe signs of anorexia, ataxia, weakness, anemia, emaciation and difficulty breathing.
The excess mortality due to Leucocytozoon in adult birds seems to occur as a result of debilitation and increased susceptibility to secondary infection.
Epidemiology
L. simondi is suspected to be a major parasite of Canadian geese in some areas, including the upper Midwestern United States and Canada. L. smithi affects turkey farms in the southeastern United States.
Host range
Bird hosts