
Family Leptospiraceae Higher classification Spirochaetaceae Phylum Spirochaetes | Order Leptospirales Scientific name Leptospira Rank Genus | |
 | ||
Similar Leptospira interrogans, Bacteria, Spirochaetes, Spirochaetaceae, Borrelia recurrentis | ||
Leptospira (Greek leptos, "fine, thin" and Latin spira, "coil") is a genus of spirochaete bacteria, including a small number of pathogenic and saprophytic species. Leptospira was first observed in 1907 in kidney tissue slices of a leptospirosis victim who was described as having died of "yellow fever."
Contents
- Leptospirosis infecci n grave causada por la bacteria leptospira
- Taxonomy
- Phylogeny
- Morphology
- Cellular structure
- Habitat
- Nutrition
- Genome
- Genotyping
- References
Leptospirosis infecci n grave causada por la bacteria leptospira
Taxonomy

Leptospira, together with the genera Leptonema and Turneria, is a member of the family Leptospiraceae. The genus Leptospira is divided into 20 species based on DNA hybridization studies.
Pathogenic Leptospira
Leptospira alstoni Smythe et al. 2013 ["Leptospira alstoni" Haake et al. 1993]Leptospira interrogans (Stimson 1907) Wenyon 1926 emend. Faine and Stallman 1982 ["Spirochaeta interrogans" Stimson 1907; "Spirochaeta nodosa" Hubener & Reiter 1916; "Spirochaeta icterohaemorrhagiae" Inada et al. 1916; "Spirochaeta icterogenes" Uhlenhuth & Fromme 1916; "Leptospira icteroides" Noguchi 1919]Leptospira kirschneri Ramadass et al. 1992Leptospira noguchii Yasuda et al. 1987Leptospira alexanderi Brenner et al. 1999Leptospira weilii Yasuda et al. 1987Leptospira borgpetersenii Yasuda et al. 1987Leptospira santarosai Yasuda et al. 1987Leptospira kmetyi Slack et al. 2009Leptospira mayottensis Bourhy et al. 2014Intermediates or opportunistic Leptospira
Leptospira inadai Yasuda et al. 1987Leptospira fainei Perolat et al. 1998Leptospira broomii Levett et al. 2006Leptospira licerasiae Matthias et al. 2009Leptospira wolffii Slack et al. 2008Non-pathogenic Leptospira
Members of Leptospira are also grouped into serovars according to their antigenic relatedness. There are currently over 200 recognized serovars. A few serovars are found in more than one species of Leptospira.

At its 2002 meeting, the Committee on the Taxonomy of Leptospira of the International Union of Microbiological Societies approved the following nomenclature for serovars of Leptospira. Genus and species must of course be italicized, with the serovar name not italicized and with an upper case first letter.
Genus species serovar Serovar_nameFor example:
Phylogeny
The currently accepted taxonomy is based on the List of Prokaryotic names with Standing in Nomenclature (LPSN) and National Center for Biotechnology Information (NCBI), and the phylogeny is based on 16S rRNA-based LTP release 123 by 'The All-Species Living Tree' Project.
Morphology
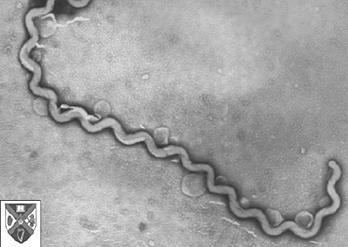
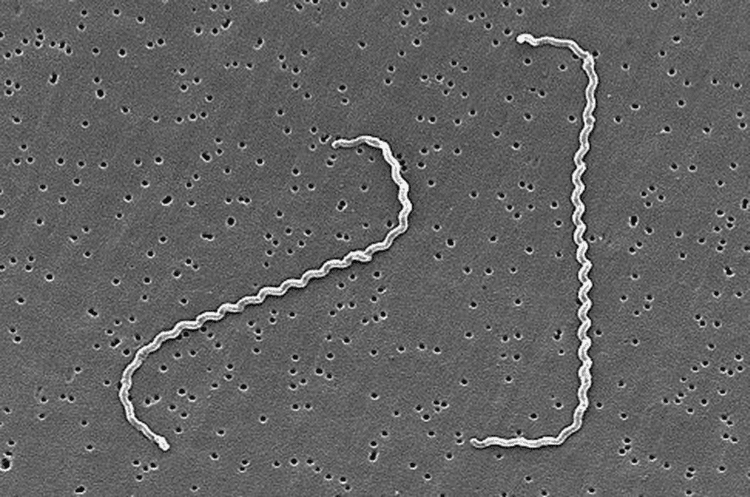
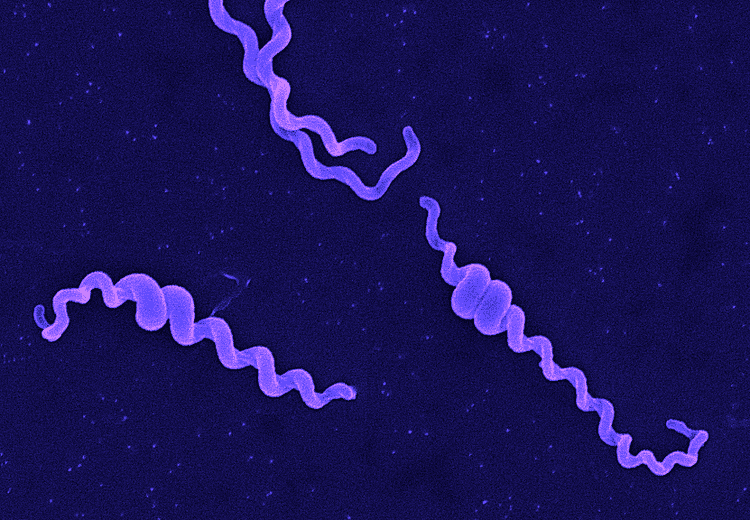
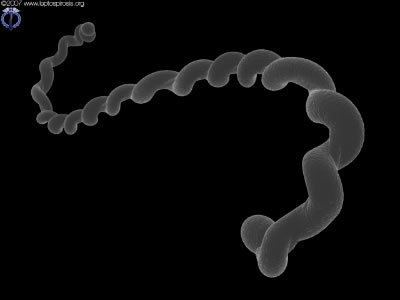
Although over 200 serotypes of Leptospira have been described, all members of the genus have similar morphology. Leptospira are spiral-shaped bacteria that are 6-20 μm long and 0.1 μm in diameter with a wavelength of about 0.5 μm. One or both ends of the spirochete are usually hooked. Because they are so thin, live Leptospira are best observed by darkfield microscopy.
The bacteria have a number of degrees of freedom; when ready to proliferate via binary fission, the bacterium noticeably bends in the place of the future split.
Cellular structure
Leptospira have a Gram-negative-like cell envelope consisting of a cytoplasmic and outer membrane. However, the peptidoglycan layer is associated with the cytoplasmic rather than the outer membrane, an arrangement that is unique to spirochetes. The two flagella of Leptospira extend from the cytoplasmic membrane at the ends of the bacteria into the periplasmic space and are necessary for the motility of Leptospira.
The outer membrane contains a variety of lipoproteins and transmembrane outer membrane proteins. As expected, the protein composition of the outer membrane differs when comparing Leptospira growing in artificial medium with Leptospira present in an infected animal. Several leptospiral outer membrane proteins have been shown to attach to the host extracellular matrix and to factor H. These proteins may be important for adhesion of Leptospira to host tissues and in resisting complement, respectively.
The outer membrane of Leptospira, like those of most other Gram-negative bacteria, contains lipopolysaccharide (LPS). Differences in the highly immunogenic LPS structure account for the numerous serovars of Leptospira. Consequently, immunity is serovar specific; current leptospiral vaccines, which consist of one or several serovars of Leptospira endemic in the population to be immunized, protect only against the serovars contained in the vaccine preparation. Leptospiral LPS has low endotoxin activity. An unusual feature of leptospiral LPS is that it activates host cells via TLR2 rather than TLR4. The unique structure of the lipid A portion of the LPS molecule may account for this observation. Finally, the LPS O antigen content of L. interrogans differs in an acutely infected versus a chronically infected animal. The role of O antigen changes in the establishment or maintenance of acute or chronic infection, if any, is unknown.
Habitat
Leptospira, both pathogenic and saprophytic, can occupy diverse environments, habitats and life cycles; these bacteria are found throughout the world, except in Antarctica. High humidity and neutral (6.9–7.4) pH are necessary for their survival in the environment, with stagnant water reservoirs—bogs, shallow lakes, ponds, puddles, etc.—being the natural habitat for the bacteria.
Nutrition
Leptospira are typically cultivated at 30 °C in Ellinghausen-McCullough-Johnson-Harris (EMJH) medium, which can be supplemented with 0.21% rabbit serum to enhance growth of fastidious strains. Growth of pathogenic Leptospira in an artificial nutrient environment such as EMJH becomes noticeable in 4–7 days; growth of saprophytic strains occur within 2–3 days. The minimal growth temperature of pathogenic species is 13–15 °C. Because the minimal growth temperature of the saprophytes is 5–10 °C, the ability of Leptospira to grow at 13 °C can be used to distinguish saprophytic from pathogenic Leptospira species. The optimal pH for growth of Leptospira is 7.2–7.6.
Leptospira are aerobes whose major carbon and energy source during in vitro growth is long-chain fatty acids, which are metabolized by beta-oxidation. Fatty acids are provided in EMJH in the form of Tween. Fatty acid molecules are bound by albumin in EMJH and are released slowly into the medium to prevent its toxic accumulation.
Like most bacteria, Leptospira require iron for growth. L. interrogans and L. biflexa have the ability to acquire iron in different forms. A TonB-dependent receptor required for utilization of the ferrous form of the iron has been identified in L. biflexa, and an ortholog of the receptor is encoded in the genome of L. interrogans. L. interrogans can also obtain iron from heme, which is bound to most of the iron in the human body. The HbpA hemin-binding protein, which may be involved in the uptake of hemin, has been identified on the surface of L. interrogans Although other pathogenic species of Leptospira and L. biflexa lack HbpA, yet another hemin-binding protein, LipL41, may account for their ability to use hemin as a source of iron. Although they do not secrete siderophores, L. biflexa and L. interrogans may be capable of obtaining iron from siderophores secreted by other microorganisms.
Genome
The genome of pathogenic Leptospira consists of two chromosomes. The size of the genomes of L. interrogans serovars Copenhageni and Lai is approximately 4.6 Mb. However, the genome of L. borgpetersenii serovar Hardjo is only 3.9 Mb in size with a large number of pseudogenes, gene fragments, and insertion sequences relative to the genomes of L. interrogans. L. interrogans and L. borgpetersenii share 2708 genes from which 656 are pathogenic specific genes. The guanine plus cytosine (GC) content is between 35% and 41%. L. borgpetersenii serovar Hardjo is usually transmitted by direct exposure to infected tissues, whereas L. interrogans is often acquired from water or soil contaminated by the urine of carrier animals harboring Leptospira in their kidneys. The high number of defective genes and insertion sequences in L. borgpetersenii Hardjo together with the poor survival outside of the host and difference in transmission patterns compared to L. interrogans suggest that L. borgpetersenii is undergoing insertion-sequence mediated genomic decay, with ongoing loss of genes necessary for survival outside of the host animal.
Genotyping
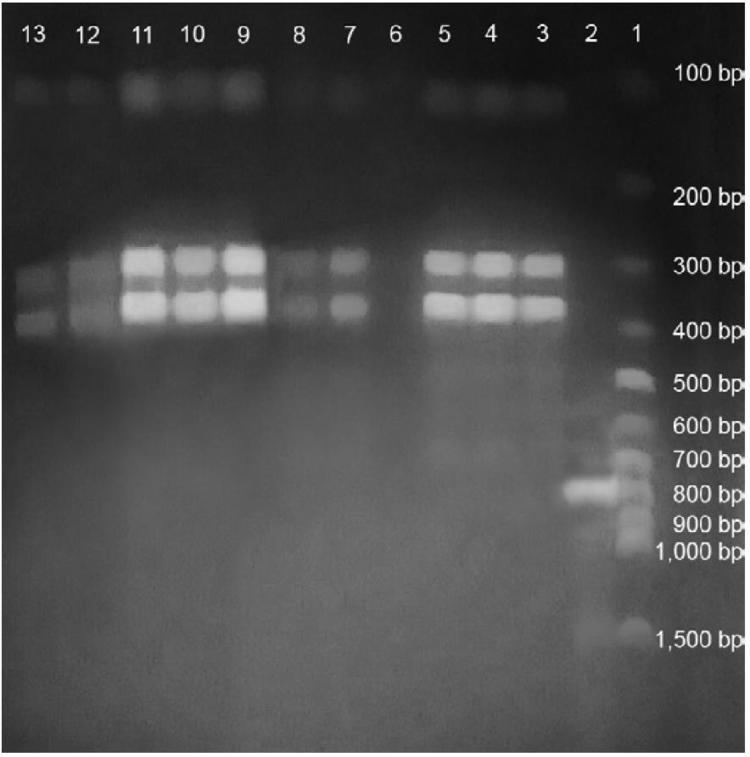
Genome sequence determination several strains of Leptospira lead to the development of multilocus VNTR (Variable Number of Tandem Repeats) typing and multilocus sequence typing (MLST) for species level identification of pathogenic Leptospira species. Both methods hold the potential to replace the highly ambiguous serotyping method currently in vogue for leptospiral strain identification.