
Species Human Entrez 3949 | Human Mouse Ensembl ENSG00000130164 | |
 | ||
External IDs OMIM: 606945 MGI: 96765 HomoloGene: 55469 GeneCards: LDLR | ||
Ldl receptor part 1 of 2 receptor mediated endocytosis
The Low-Density Lipoprotein (LDL) Receptor (LDL-R) is a mosaic protein of 839 amino acids (after removal of 21-amino acid signal peptide) that mediates the endocytosis of cholesterol-rich LDL. It is a cell-surface receptor that recognizes the apoprotein B100, which is embedded in the outer phospholipid layer of LDL particles. The receptor also recognizes the apoE protein found in chylomicron remnants and VLDL remnants (IDL). In humans, the LDL receptor protein is encoded by the LDLR gene on chromosome 19. It belongs to the Low density lipoprotein receptor gene family. It is most significantly expressed in bronchial epithelial cells and adrenal gland and cortex tissue.
Contents
- Ldl receptor part 1 of 2 receptor mediated endocytosis
- The ldl receptor part 1
- Gene
- Protein
- Mutations
- Function
- Clinical significance
- Clinical marker
- Interactive pathway map
- References
Michael S. Brown and Joseph L. Goldstein were awarded the 1985 Nobel Prize in Physiology or Medicine for their identification of LDL-R and its relation to cholesterol metabolism and familial hypercholesterolemia. The LDLR gene also contains one of 27 SNPs associated with increased risk of coronary artery disease.
The ldl receptor part 1
Gene
The LDLR gene resides on chromosome 19 at the Band 19p13.2 and is split into 18 exons. Exon 1 contains a signal sequence that localises the receptor to the endoplasmic reticulum for transport to the cell surface. Beyond this, exons 2-6 code the ligand binding region; 7-14 code the epidermal growth factor (EGF) domain; 15 codes the oligosaccharide rich region; 16 (and some of 17) code the membrane spanning region; and 18 (with the rest of 17) code the cytosolic domain.
This gene produces 6 isoforms through alternative splicing.
Protein
This protein belongs to the LDLR family and is made up of a number of functionally distinct domains, including 3 EGF-like domains, 7 LDL-R class A domains, and 6 LDL-R class B repeats.
The N-terminal domain of the LDL receptor, which is responsible for ligand binding, is composed of seven sequence repeats (~50% identical). Each repeat, referred to as a class A repeat or LDL-A, contains roughly 40 amino acids, including 6 cysteine residues that form disulfide bonds within the repeat. Additionally, each repeat has highly conserved acidic residues which it uses to coordinate a single calcium ion in an octahedral lattice. Both the disulfide bonds and calcium coordination are necessary for the structural integrity of the domain during the receptor's repeated trips to the highly acidic interior of the endosome. The exact mechanism of interaction between the class A repeats and ligand (LDL) is unknown, but it is thought that the repeats act as "grabbers" to hold the LDL. Binding of ApoB requires repeats 2-7 while binding ApoE requires only repeat 5 (thought to be the ancestral repeat).
Next to the ligand binding domain is an EGF precursor homology domain (EGFP domain). This shows approximately 30% homology with the EGF precursor gene. There are three "growth factor" repeats; A, B and C. A and B are closely linked while C is separated by the YWTD repeat region, which adopts a beta-propeller conformation (LDL-R class B domain). It is thought that this region is responsible for the pH-dependent conformational shift that causes bound LDL to be released in the endosome.
A third domain of the protein is rich in O-linked oligosaccharides but appears to show little function. Knockout experiments have confirmed that no significant loss of activity occurs without this domain. It has been speculated that the domain may have ancestrally acted as a spacer to push the receptor beyond the extracellular matrix.
The single transmembrane domain of 22 (mostly) non-polar residues crosses the plasma membrane in a single alpha helix.
The cytosolic C-terminal domain contains ~50 amino acids, including a signal sequence important for localizing the receptors to clathrin-coated pits and for triggering receptor-mediated endocytosis after binding. Portions of the cytosolic sequence have been found in other lipoprotein receptors, as well as in more distant receptor relatives.
Mutations
Mutations in the gene encoding the LDL receptor are known to cause familial hypercholesterolaemia.
There are 5 broad classes of mutation of the LDL receptor:
Function
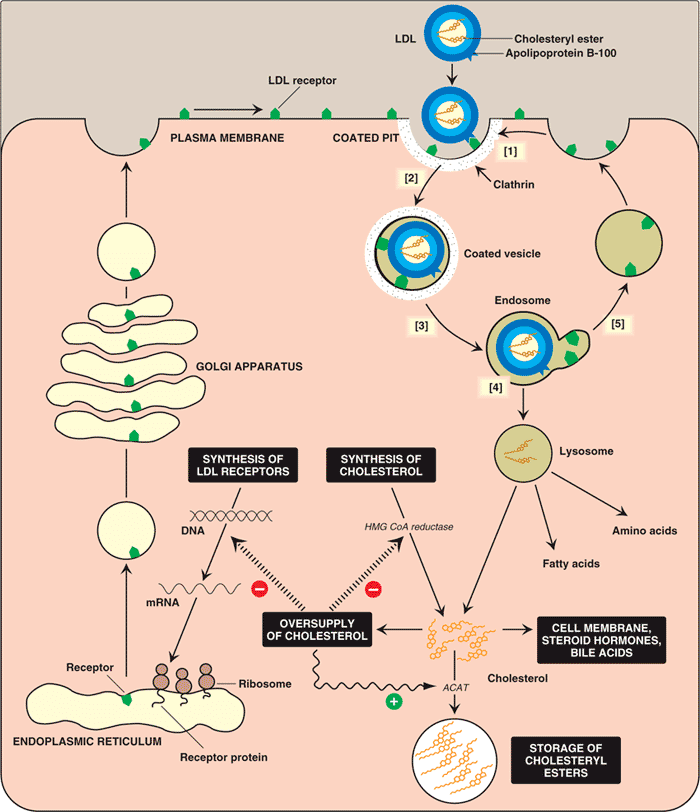
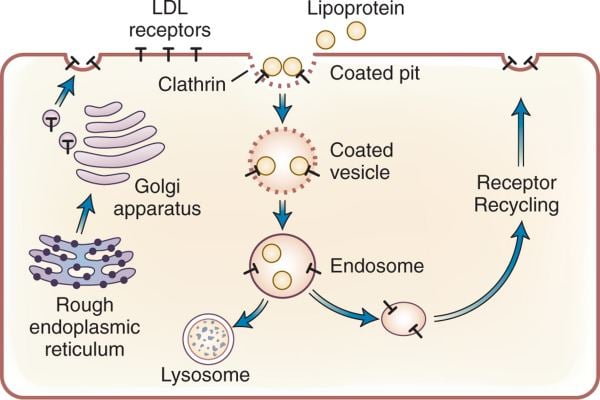
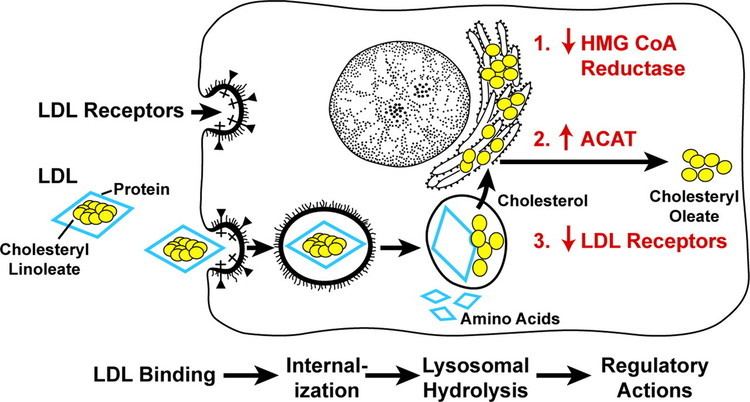
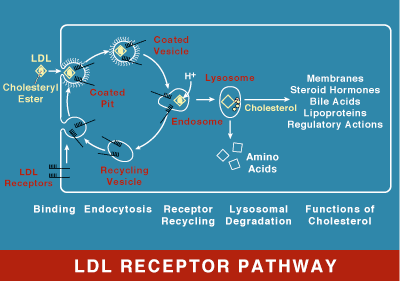
LDL receptor mediates the endocytosis of cholesterol-rich LDL and thus maintains the plasma level of LDL. This occurs in all nucleated cells, but mainly in the liver which removes ~70% of LDL from the circulation. LDL receptors are clustered in clathrin-coated pits, and coated pits pinch off from the surface to form coated endocytic vesicles that carry LDL into the cell. After internalization, the receptors dissociate from their ligands when they are exposed to lower pH in endosomes. After dissociation, the receptor folds back on itself to obtain a closed conformation and recycles to the cell surface. The rapid recycling of LDL receptors provides an efficient mechanism for delivery of cholesterol to cells. It was also reported that by association with lipoprotein in the blood, viruses such as hepatitis C virus, Flaviviridae viruses and bovine viral diarrheal virus could enter cells indirectly via LDLR-mediated endocytosis. In addition, LDLR modulation is associated with early atherosclerosis-related lymphatic dysfunction. Synthesis of receptors in the cell is regulated by the level of free intracellular cholesterol; if it is in excess for the needs of the cell then the transcription of the receptor gene will be inhibited. LDL receptors are translated by ribosomes on the endoplasmic reticulum and are modified by the Golgi apparatus before travelling in vesicles to the cell surface.
Clinical significance
In humans, LDL is directly involved in the development of atherosclerosis,which is the process responsible for the majority of cardiovascular diseases, due to accumulation of LDL-cholesterol in the blood. Hyperthyroidism may be associated with hypocholesterolaemia via upregulation of the LDL receptor, and hypothyroidism with the converse. A vast number of studies have described the relevance of LDL receptors in the pathophysiology of atherosclerosis, metabolomics syndrome, and steatohepatitis Previously, rare mutations in LDL-genes have been shown to contribute to myocardial infarction risk in individual families, whereas common variants at more than 45 loci have been associated with myocardial infarction risk in the population. When compared with non-carriers, LDLR mutation carriers had higher plasma LDL cholesterol, whereas APOA5 mutation carriers had higher plasma triglycerides. Recent evidence has connected MI risk with coding-sequence mutations at two genes functionally related to APOA5, namely lipoprotein lipase and apolipoprotein C-III. Combined, these observations suggest that, as well as LDL cholesterol, disordered metabolism of triglyceride-rich lipoproteins contributes to MI risk. Overall, LDLR has a high clinical relevance in blood lipids.
LDL is directly involved in the development of atherosclerosis, due to accumulation of LDL-cholesterol in the blood. Atherosclerosis is the process responsible for the majority of cardiovascular diseases.
Hyperthyroidism may be associated with hypocholesterolaemia via upregulation of the LDL receptor, and hypothyroidism with the converse.
Clinical marker
A multi-locus genetic risk score study based on a combination of 27 loci, including the LDLR gene, identified individuals at increased risk for both incident and recurrent coronary artery disease events, as well as an enhanced clinical benefit from statin therapy. The study was based on a community cohort study (the Malmo Diet and Cancer study) and four additional randomized controlled trials of primary prevention cohorts (JUPITER and ASCOT) and secondary prevention cohorts (CARE and PROVE IT-TIMI 22).
Interactive pathway map
Click on genes, proteins and metabolites below to link to respective articles.