
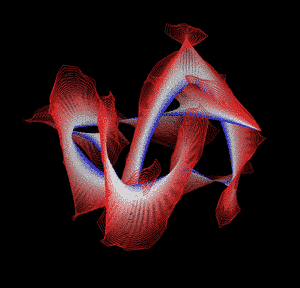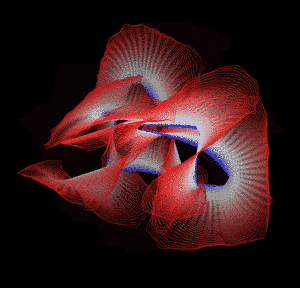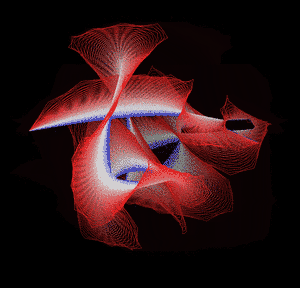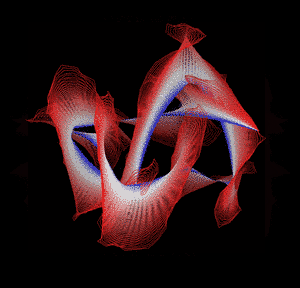 | ||
Knotted proteins are proteins whose backbones entangle themselves in a knot. One can imagine pulling a protein chain from both termini, as though pulling a string from both ends. When a knotted protein is “pulled” from both termini, it does not get disentangled. Knotted proteins are interesting because they are very rare, and their folding mechanisms and function are not well understood. Although there have been some experimental and theoretical studies that hinted to some answers, systematic answers to these questions have not been found.
Contents
- Mathematical interpretation
- Depth of the knot
- Formation of knots
- Cystine knots and slipknots
- First discoveries
- Studies about the function of the knot in a protein
- Web servers to extrapolate knotted proteins
- References
Though a number of computational methods have been developed for detecting protein knots, there are still no completely automatic methods to detect protein knots without the necessary manual intervention due to the missing residues or chain breaks in the X-ray structures or the nonstandard PDB formats.
Most of the knots discovered in proteins are deep trefoil (31) knots. Figure eight knots (42), three-twist knots (52), and Stevedore knots (61) have also been discovered.
Mathematical interpretation
Mathematically, a knot is defined as a subset of a three-dimensional points homeomorphic to a circle. According to this definition, a knot only makes sense in a closed loop. However, many strategies have been used to create an artificial closed loop. For example, if we pick a point in space at infinite distance and connect it to the N and C termini through a virtual bond, the protein can be treated as a closed loop, or stochastic methods that create random closures.
Depth of the knot
A deep knot is preserved even though the removal of a considerable number of residues from either end does not destroy the knot. The higher the number of residues that can be removed without destroying the knot, the deeper the knot.
Formation of knots
Considering how knots may be produced with a string, the folding of knotted proteins should involve first the formation of a loop, and then the threading of one terminus through the loop. This is the only topological way that the trefoil knot can be formed. For more complex knots, it is theoretically possible to have the loop to twist multiple times around itself, meaning that one end of the chain gets wrapped around at least once, and then threading to occur. It has also been observed in a theoretical study that a 6-1 knot can form by the C-terminus threading through a loop, and another loop flipping over the first loop, as well as the C-terminus threading through both the loops which have previously flipped over each other.
There have been experimental studies involving YibK and YbeA, knotted proteins containing trefoil knots. It has been established that these knotted proteins fold slowly,and that the knotting in folding is the rate limiting step. In another experimental study, a 91-residue-long protein was attached to the termini of YibK and YbeA. Attaching the protein to both termini produces a deep knot with about 125 removable residues on each terminus before the knot is destroyed. Yet it was seen that the resulting proteins could fold spontaneously. The attached proteins were shown to fold more quickly than YibK and YbeA themselves, so during folding they are expected to act as plugs at either end of YibK and YbeA. It was found that attaching the protein to the N-terminus did not alter the folding speed, but the attachment to the C-terminus slows folding down, suggesting that the threading event happens at the C-terminus.
Cystine knots and slipknots
Sometimes, post-translational knots can occur due to crosslinkings such as disulfide bonds, in which case they are called cystine knots. Proteins that contain only these are not considered knotted proteins, as the formation of these pseudo-knots in general are not different from the folding of an unknotted protein.
A slipknot in a protein is also an interesting structure. Although it is not a knot, removing a small number of residues from one of the termini may create a knot.
First discoveries
Marc L. Mansfield proposed in 1994, that there can be knots in proteins. He gave unknot scores to proteins by constructing a sphere centered at the center of mass of the alpha carbons of the backbone, with a radius twice the distance between the center of mass and the Calpha that is the farthest away from the center of mass, and by sampling two random points on the surface of the sphere. He connected the two points by tracing a geodesic on the surface of the sphere (arcs of great circles), and then connected each end of the protein chain with one of these points. Repeating this procedure a 100 times and counting the times where the knot is destroyed in the mathematical sense yields the unknot score. Human carbonic anhydrase was identified to have a low unknot score (22). Upon visually inspecting the structure, it was seen that the knot was shallow, meaning that the removal of a few residues from either end destroys the knot.
In 2000, William R. Taylor identified a deep knot in acetohydroxy acid isomeroreductase (PDB ID: 1YVE), by using an algorithm that smooths protein chains and makes knots more visible. The algorithm keeps both termini fixed, and iteratively assigns to the coordinates of each residue the average of the coordinates of the neighboring residues. It has to be made sure that the chains do not pass through each other, otherwise the crossings and therefore the knot might get destroyed. If there is no knot, the algorithm eventually produces a straight line that joins both termini.
Studies about the function of the knot in a protein
Some proposals about the function of knots have been that it might increase thermal and kinetic stability. One particular suggestion was that for the human ubiquitin hydrolase, which contains a 5-2 knot, the presence of the knot might be preventing it from being pulled into the proteasome. Because it is a deubiquitinating enzyme, it is often found in proximity of proteins soon to be degraded by proteasome, and therefore it faces the danger of being degraded itself. Therefore, the presence of the knot might be functioning as a plug that prevents it. This notion was further analyzed on other proteins like YbeA and YibK with computer simulations. The knots seem to tighten when they are pulled into a pore, and depending on the force with which they are pulled in, they either get stuck and block the pore, the likeliness of which increases with stronger pulling forces, or in the case of a small pulling force they might get disentangled as one terminus is pulled out of the knot. For deeper knots, it is more likely that the pore will be blocked, as there are too many residues that need to be pulled through the knot. In another theoretical study, it was found that the modeled knotted protein was not thermally stable, but it was kinetically stable.
Web servers to extrapolate knotted proteins
A number of web servers were available, providing convenient query services for knotted structures and analysis tools for detecting protein knots.