
 | ||
The genetic history of indigenous peoples of the Americas primarily focuses on Human Y-chromosome DNA haplogroups and Human mitochondrial DNA haplogroups. Autosomal "atDNA" markers are also used, but differ from mtDNA or Y-DNA in that they overlap significantly. The genetic pattern indicates Indigenous Amerindians experienced two very distinctive genetic episodes; first with the initial peopling of the Americas, and secondly with European colonization of the Americas. The former is the determinant factor for the number of gene lineages, zygosity mutations and founding haplotypes present in today's Indigenous Amerindian populations.
Contents
- Background
- Y DNA
- Haplogroup Q
- Haplogroup R1
- Haplogroup C P39
- mtDNA
- By autosomal DNA
- Overlaps between DNA types
- Old world genetic admixture
- Blood groups
- Genealogical testing
- References
Analyses of genetics among Amerindian and Siberian populations have been used to argue for early isolation of founding populations on Beringia and for later, more rapid migration from Siberia through Beringia into the New World. The microsatellite diversity and distributions of the Y lineage specific to South America indicates that certain Amerindian populations have been isolated since the initial colonization of the region. The Na-Dené, Inuit and Indigenous Alaskan populations exhibit Haplogroup Q-M242; however, they are distinct from other indigenous Amerindians with various mtDNA and atDNA mutations. This suggests that the peoples who first settled the northern extremes of North America and Greenland derived from later migrant populations than those who penetrated farther south in the Americas. Linguists and biologists have reached a similar conclusion based on analysis of Amerindian language groups and ABO blood group system distributions.
There is general agreement among anthropologists that the source populations for the migration into the Americas originated from an area somewhere east of the Yenisei River. The common occurrence of the mtDNA Haplogroups A, B, C, and D among eastern Asian and Amerindian populations has long been recognized, along with the presence of Haplogroup X. As a whole, the greatest frequency of the four Amerindian associated haplogroups occurs in the Altai-Baikal region of southern Siberia. Some subclades of C and D closer to the Amerindian subclades occur among Mongolian, Amur, Japanese, Korean, and Ainu populations.
Background
The Y chromosome is passed down directly from father to son; all male humans (Y chromosomes) today trace back to a single prehistoric father termed "Y-chromosomal Adam" originating in Africa. The Y chromosome spans about 60 million base pairs (the building blocks of DNA) and represents about 2 percent of the total DNA in all human cells. The original "Y chromosomal Adam"-DNA sequencing has mutated rarely over the 20,000 generations, but each time a new mutation occurs, there is a new branch in a haplogroup, resulting in a new subclade (single-nucleotide polymorphism (SNP)).
Both females and males inherit their Mitochondrial DNA (mtDNA) from only their mother. MtDNA mutations are also passed down relatively unchanged from generation to generation, so all humans share the same mtDNA-types. The logical extension of this is that all humans ultimately trace back to one woman, who is commonly referred to as Mitochondrial Eve. This line of biological inheritance, therefore, stops with each male. Consequently, Y-DNA is more commonly used by the general public for tracing genetic heritage of a direct male line.
An autosome (atDNA) is a chromosome that is not a sex chromosome – that is to say, there are an equal number of copies of the chromosome in males and females. Autosomal DNA testing is generally used to determine the "genetic percentages" of a person's ancestry from particular continents/regions or to identify the countries and "tribes" of origin on an overall basis. Genetic admixture tests arrive at these percentages by examining locations (SNPs) on the DNA where one nucleotide has "mutated" or "switched" to a different nucleotide. One way to examine the support for particular colonization routes within the American landmass is to determine whether a closer relationship between zygosity and geography is observed when "effective" geographic distances are computed along these routes, rather than along shortest-distance paths.
Y-DNA
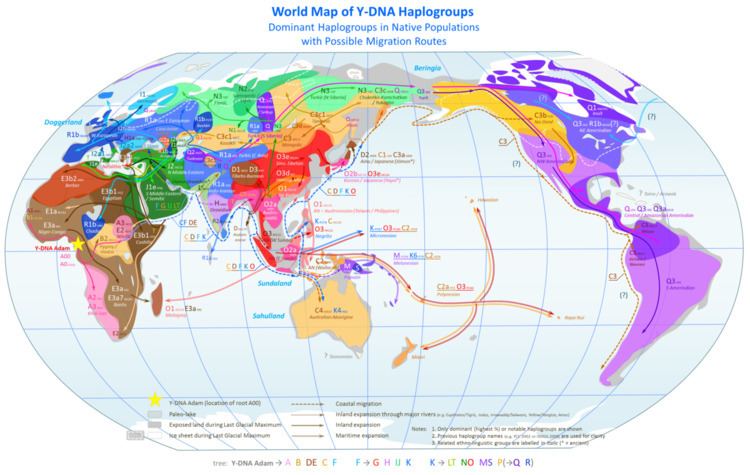
The Y chromosome consortium has established a system of defining Y-DNA haplogroups by letters A through to T, with further subdivisions using numbers and lower-case letters.
Haplogroup Q
Q-M242 (mutational name) is the defining (SNP) of Haplogroup Q (Y-DNA) (phylogenetic name). Within the Q clade, there are 14 haplogroups marked by 17 SNPs.2009 In Eurasia, haplogroup Q is found among indigenous Siberian populations, such as the modern Chukchi and Koryak peoples. In particular, two groups exhibit large concentrations of the Q-M242 mutation, the Ket (93.8%) and the Selkup (66.4%) peoples. The Ket are thought to be the only survivors of ancient wanderers living in Siberia. Their population size is very small; there are fewer than 1,500 Ket in Russia.2002 The Selkup have a slightly larger population size than the Ket, with approximately 4,250 individuals.
Starting the Paleo-Indians period, a migration to the Americas across the Bering Strait (Beringia) by a small population carrying the Q-M242 mutation took place. A member of this initial population underwent a mutation, which defines its descendant population, known by the Q-M3 (SNP) mutation. These descendants migrated all over the Americas.
Haplogroup Q-M3 is defined by the presence of the rs3894 (M3) (SNP). The Q-M3 mutation is roughly 15,000 years old as that is when the initial migration of Paleo-Indians into the Americas occurred. Q-M3 is the predominant haplotype in the Americas, at a rate of 83% in South American populations, 50% in the Na-Dené populations, and in North American Eskimo-Aleut populations at about 46%. With minimal back-migration of Q-M3 in Eurasia, the mutation likely evolved in east-Beringia, or more specifically the Seward Peninsula or western Alaskan interior. The Beringia land mass began submerging, cutting off land routes.
Since the discovery of Q-M3, several subclades of M3-bearing populations have been discovered. An example is in South America, where some populations have a high prevalence of (SNP) M19 which defines subclade Q-M19. M19 has been detected in (59%) of Amazonian Ticuna men and in (10%) of Wayuu men. Subclade M19 appears to be unique to South American Indigenous peoples, arising 5,000 to 10,000 years ago.
This suggests that population isolation and perhaps even the establishment of tribal groups began soon after migration into the South American areas. Other American subclades include Q-L54, Q-Z780, Q-MEH2, Q-SA01, and Q-M346 lineages. In Canada, two other lineages have been found. These are Q-P89.1 and Q-NWT01.
The principal-component analysis suggests a close genetic relatedness between some North American Amerindians (the Chipewyan and the Cheyenne) and certain populations of central/southern Siberia (particularly the Kets, Yakuts, Selkups, and Altays), at the resolution of major Y-chromosome haplogroups. This pattern agrees with the distribution of mtDNA haplogroup X, which is found in North America, is absent from eastern Siberia, but is present in the Altais of southern central Siberia. Similarly, the Asian populations closest to Native Americans are characterized by a predominance of lineage P-M45* and low frequencies of C-RPS4Y.
Haplogroup R1
Haplogroup R1 (Y-DNA) (specially R1b) is the second most predominant Y haplotype found among indigenous Amerindians after Q (Y-DNA). The distribution of R1 is believed by some to be associated with the re-settlement of Eurasia following the last glacial maximum. One theory put forth is that R1 entered the Americas with the initial founding population, suggesting prehistoric Amerindian immigration from Asia through Beringia and correlating mostly with the frequency of haplogroups Q-M3 and P-M45*. A second theory is that it was introduced during European colonization. R1 is very common throughout all of Eurasia except East Asia and Southeast Asia. R1 (M173) is found predominantly in North American groups like the Ojibwe (50-79%), Seminole (50%), Sioux (50%), Cherokee (47%), Dogrib (40%) and Tohono O'odham (Papago) (38%).
A study of Raghavan et al. 2013 found that autosomal evidence indicates that skeletal remain of a south-central Mongoloid Siberian child carrying R* y-dna (Mal'ta boy-1) "is basal to modern-day western Eurasians and genetically closely related to modern-day Amerindians, with no close affinity to east Asians." Sequencing of another south-central Siberian (Afontova Gora-2) revealed that "Siberian genetic signatures in modern-day Amerindians derive from a mixed ancestry of the First Americans." It is further theorized if "Mal'ta might be a missing link, a representative of the Asian population that admixed both into proto Indo-Europeans and Native Americans."
Haplogroup C-P39
Haplogroup C-M217 is mainly found in indigenous Siberians, Mongolians and Kazakhs. Haplogroup C-M217 is the most widespread and frequently occurring branch of the greater (Y-DNA) haplogroup C-M130. Haplogroup C-M217 descendant C-P39 is commonly found in today's Na-Dené speakers, with the highest frequency found among the Athabaskans at 42%. This distinct and isolated branch C-P39 includes almost all the Haplogroup C-M217 Y-chromosomes found among all indigenous peoples of the Americas. The Na-Dené groups are also unusual among indigenous peoples of the Americas in having a relatively high frequency of Q-M242 (25%).
Some researchers feel that this may indicate that the Na-Dené migration occurred from the Russian Far East after the initial Paleo-Indian colonization, but prior to modern Inuit, Inupiat and Yupik expansions.
mtDNA
Mitochondrial Eve is defined as the woman who was the matrilineal most recent common ancestor for all living humans. Mitochondrial Eve is estimated to have lived between 140,000 and 200,000 years ago, Mitochondrial Eve is the most recent common matrilineal ancestor, not the most recent common ancestor.
When studying human mitochondrial DNA (mtDNA) haplogroups, the results indicated until recently that Indigenous Amerindian haplogroups, including haplogroup X, are part of a single founding east Asian population. It also indicates that the distribution of mtDNA haplogroups and the levels of sequence divergence among linguistically similar groups were the result of multiple preceding migrations from Bering Straits populations. All Indigenous Amerindian mtDNA can be traced back to five haplogroups, A, B, C, D and X. More specifically, indigenous Amerindian mtDNA belongs to sub-haplogroups A2, B2, C1, D1, and X2a (with minor groups C4c, D2, D3, and D4h3). This suggests that 95% of Indigenous Amerindian mtDNA is descended from a minimal genetic founding female population, comprising sub-haplogroups A2, B2, C1b, C1c, C1d, and D1. The remaining 5% is composed of the X2a, D2, D3, C4, and D4h3 sub-haplogroups.
X is one of the five mtDNA haplogroups found in Indigenous Amerindian peoples. Unlike the four main American mtDNA haplogroups (A, B, C and D), X is not at all strongly associated with east Asia. Haplogroup X genetic sequences diverged about 20,000 to 30,000 years ago to give two sub-groups, X1 and X2. X2's subclade X2a occurs only at a frequency of about 3% for the total current indigenous population of the Americas. However, X2a is a major mtDNA subclade in North America; among the Algonquian peoples, it comprises up to 25% of mtDNA types. It is also present in lower percentages to the west and south of this area — among the Sioux (15%), the Nuu-chah-nulth (11%–13%), the Navajo (7%), and the Yakama (5%). Haplogroup X is more strongly present in the Near East, the Caucasus, and Mediterranean Europe. The predominant theory for sub-haplogroup X2a's appearance in North America is migration along with A, B, C, and D mtDNA groups, from a source in the Altai Mountains of central Asia.
Sequencing of the mitochondrial genome from Paleo-Eskimo remains (3,500 years old) are distinct from modern Amerindians, falling within sub-haplogroup D2a1, a group observed among today's Aleutian Islanders, the Aleut and Siberian Yupik populations. This suggests that the colonizers of the far north, and subsequently Greenland, originated from later coastal populations. Then a genetic exchange in the northern extremes introduced by the Thule people (proto-Inuit) approximately 800–1,000 years ago began. These final Pre-Columbian migrants introduced haplogroups A2a and A2b to the existing Paleo-Eskimo populations of Canada and Greenland, culminating in the modern Inuit.
A 2013 study in Nature reported that DNA found in the 24,000-year-old remains of a young boy from the archaeological Mal'ta-Buret' culture suggest that up to one-third of the indigenous Americans may have ancestry that can be traced back to western Eurasians, who may have "had a more north-easterly distribution 24,000 years ago than commonly thought" "We estimate that 14 to 38 percent of Amerindian ancestry may originate through gene flow from this ancient population," the authors wrote. Professor Kelly Graf said,
"Our findings are significant at two levels. First, it shows that Upper Paleolithic Siberians came from a cosmopolitan population of early modern humans that spread out of Africa to Europe and Central and South Asia. Second, Paleoindian skeletons like Buhl Woman with phenotypic traits atypical of modern-day indigenous Americans can be explained as having a direct historical connection to Upper Paleolithic Siberia."
A route through Beringia is seen as more likely than the Solutrean hypothesis. An abstract in a 2012 issue of the "American Journal of Physical Anthropology" states that "The similarities in ages and geographical distributions for C4c and the previously analyzed X2a lineage provide support to the scenario of a dual origin for Paleo-Indians. Taking into account that C4c is deeply rooted in the Asian portion of the mtDNA phylogeny and is indubitably of Asian origin, the finding that C4c and X2a are characterized by parallel genetic histories definitively dismisses the controversial hypothesis of an Atlantic glacial entry route into North America."
Another study, also focused on the mtDNA (that which is inherited through only the maternal line), revealed that the indigenous people of the Americas have their maternal ancestry traced back to a few founding lineages from East Asia, which would have arrived via the Bering strait. According to this study, it is probable that the ancestors of the Native Americans would have remained for a time in the region of the Bering Strait, after which there would have been a rapid movement of settling of the Americas, taking the founding lineages to South America.
According to a 2016 study, focused on mtDNA lineages, "a small population entered the Americas via a coastal route around 16.0 ka, following previous isolation in eastern Beringia for ~2.4 to 9 thousand years after separation from eastern Siberian populations. Following a rapid movement throughout the Americas, limited gene flow in South America resulted in a marked phylogeographic structure of populations, which persisted through time. All of the ancient mitochondrial lineages detected in this study were absent from modern data sets, suggesting a high extinction rate. To investigate this further, we applied a novel principal components multiple logistic regression test to Bayesian serial coalescent simulations. The analysis supported a scenario in which European colonization caused a substantial loss of pre-Columbian lineages".
By autosomal DNA
Genetic diversity and population structure in the American landmass is also done using autosomal (atDNA) micro-satellite markers genotyped; sampled from North, Central, and South America and analyzed against similar data available from other indigenous populations worldwide. The Amerindian populations show a lower genetic diversity than populations from other continental regions. Observed is a decreasing genetic diversity as geographic distance from the Bering Strait occurs, as well as a decreasing genetic similarity to Siberian populations from Alaska (the genetic entry point).
Also observed is evidence of a higher level of diversity and lower level of population structure in western South America compared to eastern South America. There is a relative lack of differentiation between Mesoamerican and Andean populations, a scenario that implies that coastal routes were easier for migrating peoples (more genetic contributors) to traverse in comparison with inland routes.
The over-all pattern that is emerging suggests that the Americas were colonized by a small number of individuals (effective size of about 70), which grew by a factor of 10 over 800 – 1000 years. The data also shows that there have been genetic exchanges between Asia, the Arctic, and Greenland since the initial peopling of the Americas.
In 2014, the autosomal DNA of a 12,500+-year-old infant from Montana was sequenced. The DNA was taken from a skeleton referred to as Anzick-1, found in close association with several Clovis artifacts. Comparisons showed strong affinities with DNA from Siberian sites, and virtually ruled out that particular individual had any close affinity with European sources (the "Solutrean hypothesis"). The DNA also showed strong affinities with all existing Amerindian populations, which indicated that all of them derive from an ancient population that lived in or near Siberia, the Upper Palaeolithic Mal'ta population.
According to an autosomal genetic study from 2012, Native Americans descend of at least three main migrant waves from East Asia. Most of it is traced back to a single ancestral population, called 'First Americans'. However, those who speak Inuit languages from the Arctic inherited almost half of their ancestry from a second East Asian migrant wave. And those who speak Na-dene, on the other hand, inherited a tenth of their ancestry from a third migrant wave. The initial settling of the Americas was followed by a rapid expansion southwards, by the coast, with little gene flow later, especially in South America. One exception to this are the Chibcha speakers, whose ancestry comes from both North and South America.
Linguistic studies have backed up genetic studies, with ancient patterns having been found between the languages spoken in Siberia and those spoken in the Americas.
Two 2015 autosomal DNA genetic studies confirmed the Siberian origins of the Natives of the Americas. However an ancient signal of shared ancestry with the Natives of Australia and Melanesia was detected among the Natives of the Amazon region. The migration coming out of Siberia would have happened 23000 years ago.
Overlaps between DNA types
Populations that have a specific combination of autosome, Y and MT-haplogroup mutations can generally be found with regional variations. Autosomes, Y mutations and mt mutations do not necessarily occur at a similar time and there are differential rates of sexual selection between the two sex chromosomes. This combined with population bottlenecks, the founder effect, mitochondrial mutations and genetic drift will alter the genetic composition of isolated populations, resulting in very distinguishable mutation patterns. (i.e. Taínos, Fuegians, Inuit, Yupik and Algonquian)
The rough overlaps between Y-DNA and mtDNA between the Americas, Circumpolar north, and Siberian indigenous populations are:
Old world genetic admixture
Interracial marriage and interracial sex and, more generally, the process of racial admixture, has its origins in prehistory. Racial mixing became widespread during European colonialism in the Age of Discovery. Genetic exchange between two populations reduces the genetic distance between the populations and is measurable in DNA patterns. During the Age of Discovery, beginning in the late 15th century, European explorers sailed the oceans, eventually reaching all the major continents. During this time Europeans contacted many populations, some of which had been relatively isolated for millennia. The genetic demographic composition of the Eastern Hemisphere has not changed significantly since the Age of Discovery. However, genetic demographics in the Western Hemisphere were radically altered by events following the voyages of Christopher Columbus. The European colonization of the Americas brought contact between peoples of Europe, Africa and Asia and the Amerindian populations. As a result, the Americas today have significant and complex multiracial populations. Many individuals who self-identify as one race exhibit genetic evidence of a multiracial ancestry.
The European conquest of Latin America beginning in the late 15th century, was initially executed by male soldiers and sailors from the Iberian Peninsula (Spain and Portugal). The new soldier-settlers fathered children with Amerindian women and later with African slaves. These mixed-race children were generally identified by the Spanish colonist and Portuguese colonist as "Castas". The subsequent North American fur trade during the 16th century brought many more European men, from France, Ireland, and Great Britain, who took North Amerindian women as wives. Their children became known as "Métis" or "Bois-Brûlés" by the French colonists and "mixed-bloods", "half-breeds" or "country-born" by the English colonists and Scottish colonists. From the second half of the 19th century to the beginning of the 20th century, new waves of immigrants from northern, eastern and southern Europe went to the Americas and consequently altered the demographics. Following World War II and subsequent worldwide migrations, the current American populations' genetic admixture can be traced to all corners of the world.
Blood groups
Prior to the 1952 confirmation of DNA as the hereditary material by Alfred Hershey and Martha Chase, scientists used blood proteins to study human genetic variation. The ABO blood group system is widely credited to have been discovered by the Austrian Karl Landsteiner, who found three different blood types in 1900. Blood groups are inherited from both parents. The ABO blood type is controlled by a single gene (the ABO gene) with three alleles: i, IA, and IB.
Research by Ludwik and Hanka Herschfeld during World War I found that the frequencies of blood groups A,B and O differed greatly from region to region. The "O" blood type (usually resulting from the absence of both A and B alleles) is very common around the world, with a rate of 63% in all human populations. Type "O" is the primary blood type among the indigenous populations of the Americas, in-particular within Central and South America populations, with a frequency of nearly 100%. In indigenous North American populations the frequency of type "A" ranges from 16% to 82%. This suggests again that the initial Amerindians evolved from an isolated population with a minimal number of individuals.
Genealogical testing
A genealogical DNA test examines the nucleotides at specific locations on a person's DNA for genetic genealogy purposes. The test results are not meant to have any medical value; they are intended only to give genealogical information. Genealogical DNA tests generally involve comparing the results of living individuals to historic populations. The general procedure for taking a genealogical DNA test involves taking a painless cheek-scraping (also known as a buccal swab) at home and mailing the sample to a genetic genealogy laboratory for testing. Genetic testing results showing specific sub-Haplogroups of Q, R1 and C3b implies that the individuals ancestry is, in whole or in-part, indigenous to the Americas. If one's mtDNA belonged to specific sub-Haplogroups of, A, B, C, D or X2a, the implication would be that the individual's ancestry is, in whole or part, indigenous to the Americas.