
 | ||
Appearance white microcrystalline powder | ||
gamma-Aminobutyric acid (γ-Aminobutyric acid) /ˈɡæmə əˈmiːnoʊbjuːˈtɪrᵻk ˈæsᵻd/ (also called GABA /ˈɡæbə/ for short) is the chief inhibitory neurotransmitter in the mammalian central nervous system. It plays the principal role in reducing neuronal excitability throughout the nervous system. In humans, GABA is also directly responsible for the regulation of muscle tone.
Contents
- Neurotransmitter
- Brain development
- Beyond the nervous system
- Structure and conformation
- History
- Biosynthesis
- Catabolism
- Pharmacology
- GABAergic drugs
- In plants
- References
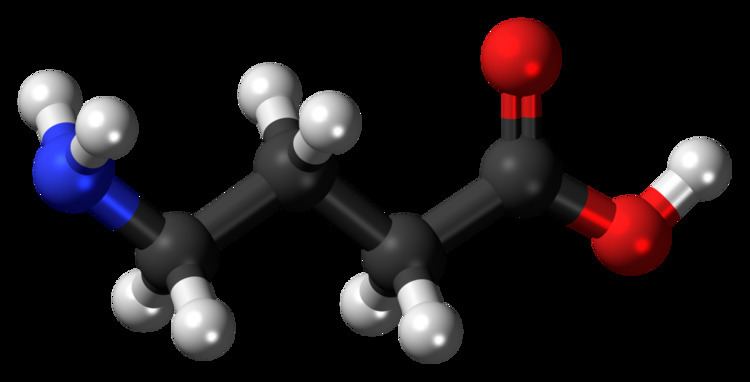
Although in chemical terms it is an amino acid (as it has both a primary amine and a carboxylic acid functional group), GABA is rarely referred to as such in the scientific or medical community. By convention the term "amino acid", when used without a qualifier, refers specifically to an alpha amino acid. GABA is not an alpha amino acid, meaning the amino group is not attached to the alpha carbon so it is not incorporated into proteins.
Neurotransmitter
In vertebrates, GABA acts at inhibitory synapses in the brain by binding to specific transmembrane receptors in the plasma membrane of both pre- and postsynaptic neuronal processes. This binding causes the opening of ion channels to allow the flow of either negatively charged chloride ions into the cell or positively charged potassium ions out of the cell. This action results in a negative change in the transmembrane potential, usually causing hyperpolarization. Two general classes of GABA receptor are known:
Neurons that produce GABA as their output are called GABAergic neurons, and have chiefly inhibitory action at receptors in the adult vertebrate. Medium spiny cells are a typical example of inhibitory central nervous system GABAergic cells. In contrast, GABA exhibits both excitatory and inhibitory actions in insects, mediating muscle activation at synapses between nerves and muscle cells, and also the stimulation of certain glands. In mammals, some GABAergic neurons, such as chandelier cells, are also able to excite their glutamatergic counterparts.
GABAA receptors are ligand-activated chloride channels: when activated by GABA, they allow the flow of chloride ions across the membrane of the cell. Whether this chloride flow is depolarizing (makes the voltage across the cell's membrane less negative), shunting (has no effect on the cell's membrane potential), or inhibitory/hyperpolarizing (makes the cell's membrane more negative) depends on the direction of the flow of chloride. When net chloride flows out of the cell, GABA is depolarising; when chloride flows into the cell, GABA is inhibitory or hyperpolarizing. When the net flow of chloride is close to zero, the action of GABA is shunting. Shunting inhibition has no direct effect on the membrane potential of the cell; however, it reduces the effect of any coincident synaptic input by reducing the electrical resistance of the cell's membrane. Shunting inhibition can "override" the excitatory effect of depolarising GABA, resulting in overall inhibition even if the membrane potential becomes less negative. It was thought that a developmental switch in the molecular machinery controlling concentration of chloride inside the cell changes the functional role of GABA between neonatal and adult stages. As the brain develops into adulthood, GABA's role changes from excitatory to inhibitory. However, this theory of excitatory GABA in developing brain has been questioned and subsequent studies in live neonatal rodents have directly shown GABA to be inhibitory in its action (see next section).
Brain development
While GABA is an inhibitory transmitter in the mature brain, its actions were thought to be primarily excitatory in the developing brain. The gradient of chloride was reported to be reversed in immature neurons, with its reversal potential higher than the resting membrane potential of the cell; activation of a GABA-A receptor thus leads to efflux of Cl− ions from the cell (that is, a depolarizing current). The differential gradient of chloride in immature neurons was shown to be primarily due to the higher concentration of NKCC1 co-transporters relative to KCC2 co-transporters in immature cells. GABA itself is partially responsible for orchestrating the maturation of ion pumps. GABAergic interneurons mature faster in the hippocampus and the GABA signalling machinery appears earlier than glutamatergic transmission. Thus, GABA was considered the major excitatory neurotransmitter in many regions of the brain before the maturation of glutamatergic synapses.
However, this theory has been questioned based on results showing that in brain slices of immature mice incubated in artificial cerebrospinal fluid (ACSF) (modified in a way that takes into account the normal composition of the neuronal milieu in sucklings by adding an energy substrate alternative to glucose, beta-hydroxybutyrate) GABA action shifts from excitatory to inhibitory mode.
This effect has been later repeated when other energy substrates, pyruvate and lactate, supplemented glucose in the slices' media. Later investigations of pyruvate and lactate metabolism found that the original results were not due to energy source issues but to changes in pH resulting from the substrates acting as "weak acids". These arguments were later rebutted by further findings showing that changes in pH even greater than that caused by energy substrates do not affect the GABA-shift described in the presence of energy substrate-fortified ACSF and that the mode of action of beta-hydroxybutyrate, pyruvate and lactate (assessed by measurement NAD(P)H and oxygen utilization) was energy metabolism-related. The true nature GABA effect in the developing brain has remained elusive until 2015, when the first study to directly show GABA action in live rodent brain has reported GABA to not be excitatory in its effect even though it slightly depolarised some neurons, confirming the dominance of shunting inhibition. In 2016, another study in live developing brains using optogenetics have shown GABA to be inhibitory, with GABAergic synapse activation leading to a reduction of network activity. That study has also shown that the same technique when used in brain slices instead records excitatory GABA effects, confirming the fact that excitatory GABA is most likely an artefact of in-vitro slice recordings.
In the developmental stages preceding the formation of synaptic contacts, GABA is synthesized by neurons and acts both as an autocrine (acting on the same cell) and paracrine (acting on nearby cells) signalling mediator. The ganglionic eminences also contribute greatly to building up the GABAergic cortical cell population.
GABA regulates the proliferation of neural progenitor cells the migration and differentiation the elongation of neurites and the formation of synapses.
GABA also regulates the growth of embryonic and neural stem cells. GABA can influence the development of neural progenitor cells via brain-derived neurotrophic factor (BDNF) expression. GABA activates the GABAA receptor, causing cell cycle arrest in the S-phase, limiting growth.
Beyond the nervous system
GABAergic mechanisms have been demonstrated in various peripheral tissues and organs including, but not restricted to, the intestine, stomach, pancreas, Fallopian tube, uterus, ovary, testis, kidney, urinary bladder, lung, and liver.
In 2007, an excitatory GABAergic system was described in the airway epithelium. The system activates following exposure to allergens and might participate in the mechanisms of asthma. GABAergic systems have also been found in the testis and in the eye lens.
GABA occurs in plants.
Structure and conformation
GABA is found mostly as a zwitterion, that is, with the carboxy group deprotonated and the amino group protonated. Its conformation depends on its environment. In the gas phase, a highly folded conformation is strongly favored because of the electrostatic attraction between the two functional groups. The stabilization is about 50 kcal/mol, according to quantum chemistry calculations. In the solid state, a more extended conformation is found, with a trans conformation at the amino end and a gauche conformation at the carboxyl end. This is due to the packing interactions with the neighboring molecules. In solution, five different conformations, some folded and some extended, are found as a result of solvation effects. The conformational flexibility of GABA is important for its biological function, as it has been found to bind to different receptors with different conformations. Many GABA analogues with pharmaceutical applications have more rigid structures in order to control the binding better.
History
In 1883, gamma-aminobutyric acid (GABA) was first synthesized, and it was first known only as a plant and microbe metabolic product.
In 1950, GABA was discovered to also be an integral part of the mammalian central nervous system.
Biosynthesis
Exogenous GABA does not penetrate the blood–brain barrier; it is synthesized in the brain. It is synthesized from glutamate using the enzyme glutamate decarboxylase (GAD) and pyridoxal phosphate (which is the active form of vitamin B6) as a cofactor. This process converts glutamate, the principal excitatory neurotransmitter, into the principal inhibitory neurotransmitter (GABA).
GABA is converted back to glutamate by a metabolic pathway called the GABA shunt.
Catabolism
GABA transaminase enzyme catalyzes the conversion of 4-aminobutanoic acid (GABA) and 2-oxoglutarate (α-ketoglutarate) into succinic semialdehyde and glutamate. Succinic semialdehyde is then oxidized into succinic acid by succinic semialdehyde dehydrogenase and as such enters the citric acid cycle as a usable source of energy.
Pharmacology
Drugs that act as allosteric modulators of GABA receptors (known as GABA analogues or GABAergic drugs) or increase the available amount of GABA typically have relaxing, anti-anxiety, and anti-convulsive effects. Many of the substances below are known to cause anterograde amnesia and retrograde amnesia.
In general, GABA does not cross the blood–brain barrier, although certain areas of the brain that have no effective blood–brain barrier, such as the periventricular nucleus, can be reached by drugs such as systemically injected GABA. At least one study suggests that orally administered GABA increases the amount of human growth hormone (HGH). GABA directly injected to the brain has been reported to have both stimulatory and inhibitory effects on the production of growth hormone, depending on the physiology of the individual. Certain pro-drugs of GABA (ex. picamilon) have been developed to permeate the blood–brain barrier, then separate into GABA and the carrier molecule once inside the brain. This allows for a direct increase of GABA levels throughout all areas of the brain, in a manner following the distribution pattern of the pro-drug prior to metabolism.
GABA enhanced the catabolism of serotonin into N-acetylserotonin (the precursor of melatonin) in rats. It is thus suspected that GABA is involved in the synthesis of melatonin and thus might exert regulatory effects on sleep and reproductive functions.
GABAergic drugs
In plants
GABA is also found in plants. It is the most abundant amino acid in the apoplast of tomatoes. It has also a role in cell signalling in plants.