
Kingdom Animalia Scientific name Eurypterida Phylum Arthropoda | Subphylum Chelicerata Higher classification Merostomata Rank Order | |
 | ||
Lower classifications Pterygotus, Pentecopterus, Pagea | ||
Ark survival evolved eurypterid farming s4e36 the center map gameplay
Eurypterids (sea scorpions) are an extinct group of arthropods related to arachnids that include the largest known arthropods to have ever lived. They are members of the extinct order Eurypterida (Chelicerata); which is the most diverse Paleozoic chelicerate order in terms of species. The name Eurypterida comes from the Greek words eury- (meaning "broad" or "wide") and pteron (meaning "wing"). This name was chosen due to the pair of wide swimming appendages on the first fossil eurypterids discovered. The largest, such as Jaekelopterus, reached 2.5 metres (8 ft 2 in) in length, but most species were less than 20 centimetres (8 in). They were formidable predators that thrived in warm shallow water, in both seas and lakes, from the mid Ordovician to late Permian (470 to 248 million years ago).
Contents
- Ark survival evolved eurypterid farming s4e36 the center map gameplay
- Body structure
- Relationships with other groups
- List of families and genera
- Phylogeny
- References

Although informally called "sea scorpions", only the earliest ones were marine (later ones lived in brackish or freshwater), and they were not true scorpions. According to theory, the move from the sea to fresh water had probably occurred by the Pennsylvanian subperiod. Eurypterids are believed to have undergone ecdysis, making their significance in ecosystems difficult to assess, because it can be difficult to tell a fossil moult from a true fossil carcass. They became extinct during the Permian–Triassic extinction event or sometime before the event 252.17 million years ago. Their fossils have a near global distribution.

About two dozen families of eurypterids are known. Perhaps the best-known genus of eurypterid is Eurypterus, of which around 16 fossil species are known. The genus Eurypterus was described in 1825 by James Ellsworth De Kay, a zoologist. He recognized the arthropod nature of the first-ever described eurypterid specimen, found by Dr. S. L. Mitchill. In 1984, that species, Eurypterus remipes was named the state fossil of New York.

Pentecopterus decorahensis, which lived as early as 467.3 million years ago, is the oldest described eurypterid; with an estimated length of 1.83 metres (6 ft 0 in), it has been described as "the first real big predator".

Body structure
Eurypterids have been formally described as follows:
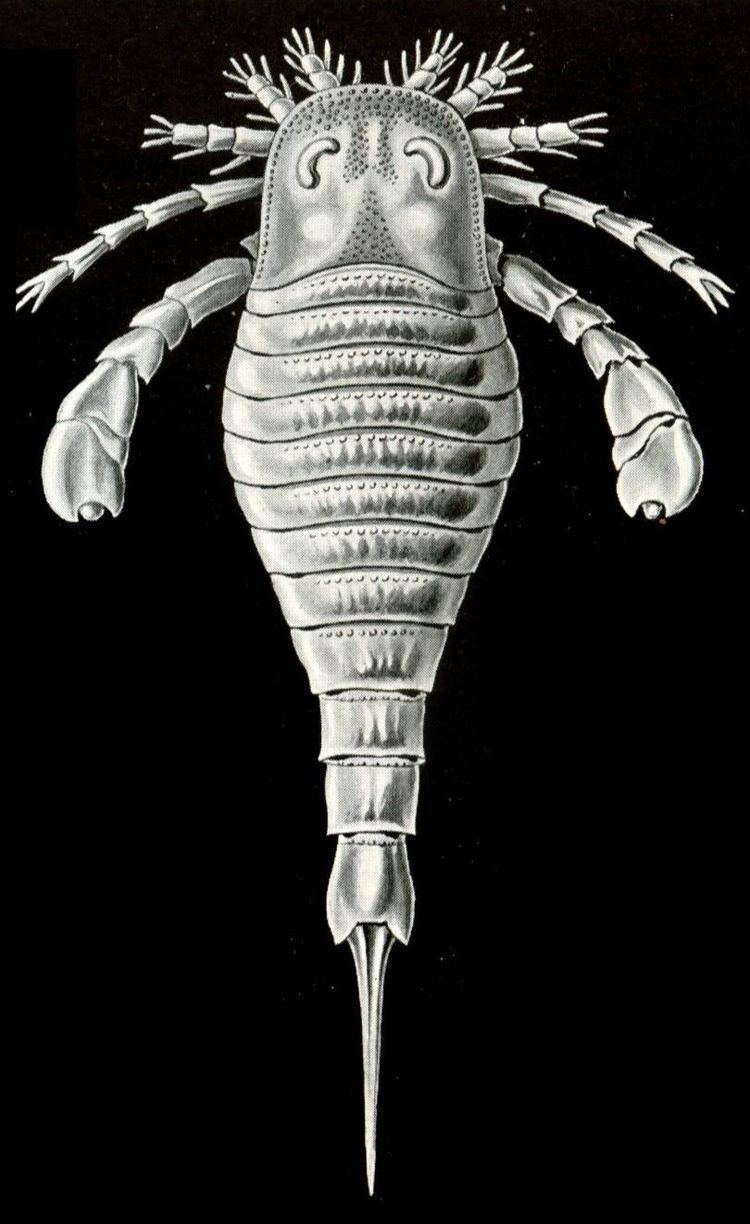
Small to very large merostomes with elongate lanceolate, rarely trilobed body; prosoma [head] of moderate size; opisthosoma [body] with 12 moveable segments and styliform to spatulate telson [tail], with division commonly into 7-segmented preabdomen and 5-segmented postabdomen; prosomal [head] appendages 6, comprising 3-jointed chelicerae, walking legs, the last pair commonly transformed into swimming legs. Mouth central, bordered posteriorly by endostoma and metastoma. Operculum with median genital appendage, abdominal appendages plate-shaped with nonlaminate gills. Ordovician-Permian.

The typical eurypterid had a large, flat, semicircular carapace, followed by a jointed section, and finally a tapering, flexible tail, most ending with a long spine at the end (Pterygotus, though, had a large flat tail, possibly with a smaller spine). Behind the head, eurypterids had twelve body segments, each of which was formed from a dorsal plate (tergite) and a ventral plate (sternite). The tail, known as the telson, is spiked in most eurypterids (as in modern scorpions) and in some species it may have been used to inject venom, but there is no evidence that any eurypterids were venomous. Most eurypterids have paddles toward the end of the carapace and beyond, which were used to propel themselves through water. The suborder Stylonurina had walking legs instead of paddles. Some argue that the paddles were also used for digging. Underneath, in addition to the pair of swimming appendages, the creature had four pairs of jointed legs for walking, and two claws at the front, chelicerae, which were enlarged in pterygotids. The walking legs had odd hairs, similar to modern-day crabs. Other features, common to ancient and modern arthropods of this type, include one pair of compound eyes and a pair of smaller eye spots, called ocelli, located between the other, larger, pair of eyes.
Many eurypterids had legs large and long enough to do more than allow them to crawl over the sea bottom; a number of species (particularly hibbertopterids) had large stout legs, and were probably capable of terrestrial locomotion (like terrestrial crabs today). Studies of what are believed to be fossil trackways indicate that eurypterids used in-phase, hexapodous (six-legged) and octopodous (eight-legged) gaits. Some species may have been amphibious, emerging onto land for at least part of their life cycle; they may have been capable of breathing both in water and in air. A predatory arthropod known as Protichnites, whose traces are found in Cambrian strata dating from 510 million years ago, is a possible stem group eurypterid, and is among the first evidence of animals on land.
The largest sea-scorpion and the largest known arthropod ever to have lived is Jaekelopterus rhenaniae, supported by the finding of a 46 cm (18 in) long claw in 2007, indicating a body length of 2.5 m (8 ft 2 in).
Relationships with other groups
Eurypterids have traditionally been regarded as close relatives of horseshoe crabs (Xiphosura), together forming a group called Merostomata. Subsequent studies placed eurypterids closer to the arachnids in a group called Metastomata.
There has also been a prevailing idea that eurypterids are closely related to scorpions, which they resemble. This hypothesis is reflected in the common name "sea scorpion". More recently it has been recognised that a little-known, extinct group called Chasmataspida also shares features with Eurypterida, and the two groups were sometimes confused with one another.
A recent summary of the relationships between arachnids and their relatives recognised Eurypterida, Xiphosura and Arachnida as three major groups, but was not able to resolve the phylogenetic relationship of any shared details between them. Another suggested the eurypterids were sister group to the chasmataspids, with these two groups in turn sister group to the horseshoe crabs.
List of families and genera
There are 246 valid species of eurypterids as of 2011. All of them are extinct. They are grouped into the following:
The trace fossil trackways produced by eurypterids are placed in the ichnogenus Palmichnium.
Phylogeny
The cladogram presented here is simplified from a study by Tetlie. The most important phylogenetic breakdown is based on the two major innovations that characterise the evolution of the eurypterids. The most important was the transformation of the posteriormost prosomal appendage into a swimming paddle (as found in the clade Eurypterina). The second innovation was the enlargement of the chelicerae, (as found in the family Pterygotidae), allowing these appendages to be used for active prey capture.
Seventy-five percent of eurypterid species are eurypterines; this represents 99% of specimens. The superfamily Pterygotioidea is the most species-rich clade, with 56 species, followed by the Adelophthalmoidea with 43 species; as sister taxa, they comprise the most derived eurypterids. Pterygotioidea includes the pterygotids, which are the only eurypterids to have a cosmopolitan distribution. This clade is one of the best supported within the eurypterids.
It has been suggested that the development of dermal armour in certain groups of jawless vertebrates (such as the Heterostraci and the Osteostraci) is in response to predation pressure by increasingly sophisticated eurypterid predators (specifically the pterygotids) although this has yet to be verified by detailed analysis. An increase in fish diversity is tied to a decline in eurypterid diversity in the Lower Devonian, although it is not thought that this represents competitive replacement; in fact, this is rare in the fossil record.