
 | ||
Enhancer RNAs (eRNAs) represent a class of relatively short non-coding RNA molecules (50-2000 nucleotides) transcribed from the DNA sequence of enhancer regions. They were first detected in 2010 through the use of genome-wide techniques such as RNA-seq and ChIP-seq. eRNAs can be subdivided into two main classes: 1D eRNAs and 2D eRNAs, which differ primarily in terms of their size, polyadenylation state, and transcriptional directionality. The expression of a given eRNA seems to correlate with the activity of its corresponding enhancer in a context-dependent fashion. Increasing evidence suggests that eRNAs actively play a role in transcriptional regulation in cis and in trans, and while their mechanisms of action remain unclear, a few models have been proposed.
Contents
Discovery
Enhancers as sites of extragenic transcription were initially discovered in genome-wide studies that identified enhancers as common regions of RNA polymerase II (RNA pol II) binding and non-coding RNA transcription. The level of RNA pol II-enhancer interaction and RNA transcript formation were found to be highly variable among these initial studies. Using explicit chromatin signature peaks, a significant proportion (~70%) of extragenic RNA Pol II transcription start sites were found to overlap enhancer sites in murine macrophages. Out of 12,000 neuronal enhancers in the mouse genome, almost 25% of the sites were found to bind RNA Pol II and generate transcripts. These eRNAs, unlike messenger RNAs (mRNAs), lacked modification by polyadenylation, were generally short and non-coding, and were bidirectionally transcribed. Later studies revealed the transcription of another type of eRNAs, generated through unidirectional transcription, that were longer and contained a poly A tail. Furthermore, eRNA levels were correlated with mRNA levels of nearby genes, suggesting the potential regulatory and functional role of these non-coding enhancer RNA molecules.
Summary
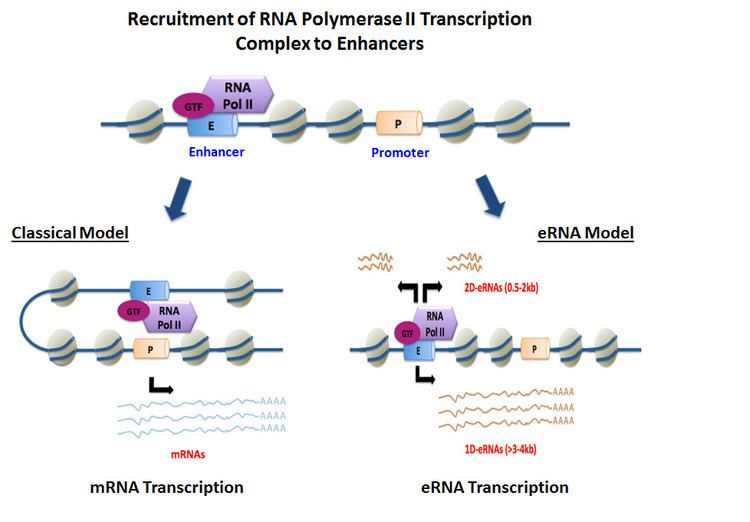
eRNAs are transcribed from DNA sequences upstream and downstream of extragenic enhancer regions. Previously, several model enhancers have demonstrated the capability to directly recruit RNA Pol II and general transcription factors and form the pre-initiation complex (PIC) prior to the start of transcription at the promoter of genes. In certain cell types, activated enhancers have demonstrated the ability to both recruit RNA Pol II and also provide a template for active transcription of their local sequences.>
Depending on the directionality of transcription, enhancer regions generate two different types of non-coding transcripts, 1D-eRNAs and 2D-eRNAs. The nature of the pre-initiation complex and specific transcription factors recruited to the enhancer may control the type of eRNAs generated. After transcription, the majority of eRNAs remain in the nucleus. In general, eRNAs are very unstable and actively degraded by the nuclear exosome. Not all enhancers are transcribed, with non-transcribed enhancers greatly outnumbering the transcribed ones in the order of magnitude of dozens of thousands in every given cell type.
1D eRNAs
In most cases, unidirectional transcription of enhancer regions generates long (>4kb) and polyadenylated eRNAs. Enhancers that generate polyA+ eRNAs have a lower H3K4me1/me3 ratio in their chromatin signature than 2D-eRNAs. PolyA+ eRNAs are distinct from long multiexonic poly transcripts (meRNAs) that are generated by transcription initiation at intragenic enhancers. These long non-coding RNAs, which accurately reflect the host gene’s structure except for the alternative first exon, display poor coding potential. As a result, polyA+ 1D-eRNAs may represent a mixed group of true enhancer-templated RNAs and multiexonic RNAs.
2D eRNAs
Bidirectional transcription at enhancer sites generates comparatively shorter (0.5-2kb) and non-polyadenylated eRNAs. Enhancers that generate polyA- eRNAs have a chromatin signature with a higher H3K4me1/me3 ratio than 1D-eRNAs. In general, enhancer transcription and production of bidirectional eRNAs demonstrate a strong correlation of enhancer activity on gene transcription.
Proposed mechanisms of function
The notions that not all enhancers are transcribed at the same time and that eRNA transcription correlates with enhancer-specific activity support the idea that individual eRNAs carry distinct and relevant biological functions. However, there is still no consensus on the functional significance of eRNAs. Furthermore, eRNAs can easily be degraded through exosomes and nonsense-mediated decay, which limits their potential as important transcriptional regulators. To date, four main models of eRNA function have been proposed, each supported by different lines of experimental evidence.
Transcriptional Noise
Since multiple studies have shown that RNA Pol II can be found at a very large number of extragenic regions, it is possible that eRNAs simply represent the product of random “leaky” transcription and carry no functional significance. The non-specific activity of RNA Pol II would therefore allow extragenic transcriptional noise at sites where chromatin is already in an open and transcriptionally competent state. This would explain even tissue-specific eRNA expression as open sites are tissue-specific as well.
Transcription-dependent effects
RNA Pol II-mediated gene transcription induces a local opening of chromatin state through the recruitment of histone acetyltransferases and other histone modifiers that promote euchromatin formation. It was proposed that the presence of these enzymes could also induce an opening of chromatin at enhancer regions, which are usually present at distant locations but can be recruited to target genes through looping of DNA. In this model, eRNAs are therefore expressed in response to RNA Pol II transcription and therefore carry no biological function.
Functional activity in cis
While the two previous models implied that eRNAs were not functionally relevant, this mechanism states that eRNAs are functional molecules that exhibit cis activity. In this model, eRNAs can locally recruit regulatory proteins at their own site of synthesis. Supporting this hypothesis, transcripts originating from enhancers upstream of the Cyclin D1 gene are thought to serve as adaptors for the recruitment of histone acetyltransferases. It was found that depletion of these eRNAs led to Cyclin D1 transcriptional silencing.
Functional activity in trans
The last model involves transcriptional regulation by eRNAs at distant chromosomal locations. Through the differential recruitment of protein complexes, eRNAs can affect the transcriptional competency of specific loci. Evf-2 represents a good example of such trans regulatory eRNA as it can induce the expression of Dlx2, which in turn can increase the activity of the Dlx5 and Dlx6 enhancers. It must be noted that trans-acting eRNAs might also be working in cis, and vice versa.
Experimental detection
The detection of eRNAs is fairly recent (2010) and has been made possible through the use of genome-wide investigation techniques such as RNA sequencing (RNA-seq) and chromatin immunoprecipitation-sequencing (ChIP-seq). RNA-seq permits the direct identification of eRNAs by matching the detected transcript to the corresponding enhancer sequence through bioinformatic analyses. ChIP-seq represents a less direct way to assess enhancer transcription but can also provide crucial information as specific chromatin marks are associated with active enhancers. Although some data remain controversial, the consensus in the literature is that the best combination of histone post-translational modifications at active enhancers is made of H2AZ, H3K27ac, and a high ratio of H3K4me1 over H3K4me3. ChIP experiments can also be conducted with antibodies that recognize RNA Pol II, which can be found at sites of active transcription. The experimental detection of eRNAs is complicated by their low endogenous stability conferred by exosome degradation and nonsense-mediated decay. Nonetheless, the fact that eRNAs tend to be expressed from active enhancers might make their detection a useful tool to distinguish between active and inactive enhancers.
Implications in development and disease
Evidence that eRNAs cause downstream effects on the efficiency of enhancer activation and gene transcription suggests its functional capabilities and potential importance. The transcription factor p53 has been demonstrated to bind enhancer regions and generate eRNAs in a p53-dependent manner. In cancer, p53 plays a central role in tumor suppression as mutations of the gene are shown to appear in 50% of tumors. These p53-bound enhancer regions (p53BERs) are shown to interact with multiple local and distal gene targets involved in cell proliferation and survival. Furthermore eRNAs generated by the activation of p53BERs are shown to be required for efficient transcription of the p53 target genes, indicating the likely important regulatory role of eRNAs in tumor suppression and cancer.
Variations in enhancers have been implicated in human disease but a therapeutic approach to manipulate enhancer activity is currently not possible. With the emergence of eRNAs as important components in enhancer activity, powerful therapeutic tools such as RNAi may provide promising routes to target disruption of gene expression.