
 | ||
Most animal cells take up portions of their surface plasma membranes in a process called endocytosis. The main route of endocytosis is the coated pit, which buds into a cell to form a cytoplasmic vesicle — a clathrin-coated vesicle. The membrane so internalised is processed in a series of intracellular organelles, which include endosomes and lysosomes. Some of this membrane is returned to the cell surface by a process called exocytosis. The whole cycle of endocytosis plus exocytosis is known as the endocytic cycle.
Contents
The endocytic cycle is crucial for the survival of individual cells and multicellular organisms.
LDL metabolism
Low-density lipoprotein (LDL) originates in the liver and is transported around an animal by the blood stream. From there it is taken up by other cells, such as fibroblasts, and degraded: this provides a source of cholesterol for the growth of these other cells. LDL in the blood binds to LDL receptors on the surface of fibroblasts; these receptors concentrate in coated pits (they are about 200x as concentrated here as along the rest of the cell’s plasma membrane) and are internalised when the pit becomes a coated vesicle. The itinerary of the LDL receptor inside the cell is complex, but it spends little time there. Within a fraction of a minute, it has released its LDL cargo and is returned to the cell surface by exocytosis. It is now ready for another round of LDL clearance.
High levels of LDL in the blood are observed in atherosclerosis and associated with the disease; the endocytic cycle reduces LDL through consuming it. This may or may not be useful in regulating increased levels of LDL, although it may have limitations, or not increase at all in the presence of extra LDL molecules.
Transferrin receptor
Transferrin is a plasma protein that is able to combine with iron ions: It is the vehicle with which iron is carried around the body. Free ferric ions are toxic; but cells need iron to build many of their proteins including cytochromes and hemoglobin. Ferric ions are carried in the blood tightly bound to transferrin as ferritransferrin. Dividing cells, which need the iron, gain it by binding the ferritransferrin to transferrin receptors on their surfaces. These receptors also have a high affinity for coated pits. Like the LDL receptor, the transferrin receptor is internalised into a coated vesicle. The iron is released inside the cell and the receptor returned to the cell surface. The route this receptor takes inside the cell appears to be different from that taken by the LDL receptor, because it takes about 10 minutes before it is exocytosed.
Synapse function
Impulses between neurons are transmitted by the release of neurotransmitters at the junction between the two cells, a region called a synapse. This release is effected by exocytosis at the presynaptic terminal. A vesicle full of transmitter, acetylcholine (for example), in the presynaptic terminal fuses with its neighbouring plasma membrane and thereby releases a burst of acetylcholine into the synaptic space. The acetylcholine is rapidly degraded here, but before this happens it activates acetylcholine receptors on the postsynaptic terminal and triggers an electrical impulse in that cell. The membrane added to the presynaptic terminal is recovered by endocytosis and recycled to form fresh vesicles full of neurotransmitter, ready for another cycle of postsynaptic excitation.
Thus, the function of the nervous system is dependent on this endocytic cycle. An example of this dependence is found in fruit flies. A key protein required for endocytosis is dynamin: It assists in budding a coated pit into a cell to form a coated vesicle. Mutations in the dynamin gene in which the activity of the dynamin protein is lost at above-normal temperatures (for the fly) exist: These are called temperature-sensitive mutations. Such mutant flies have the property that, when the fly is brought from its normal 22°C to 30°C, the dynamin function is lost. However, when the flies are cooled to 22°C, it is regained. In other words, in these mutant flies, the endocytic cycle can be turned off at 30°C, and turned back on by cooling. What one observes is that, within seconds of warming to 30°C, the fruit flies become paralysed: They drop out of the air and appear near-dead. On cooling, they slowly get up, flutter their wings and fly away. The endocytic cycle has been temporarily suspended with drastic effects.
Non-polarised cells
Animal cells, such as fibroblasts, as grown in culture in the laboratory are usually stationary; they grow and divide, but rarely move about. They have a normal endocytic cycle: coated pits 'bud in' from all over the cell’s surface in a random fashion and the returned membrane is exocytosed at the cell’s surface, also at random.
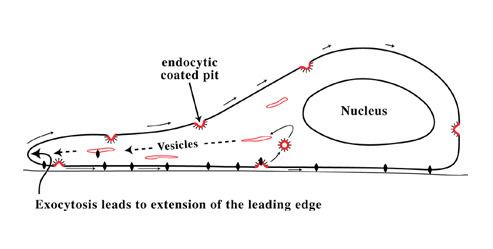
Polarised cells
Moving cells, such as fibroblasts, are arranged quite differently. Endocytosis by coated pits occurs, as in stationary cells, at random. But in motile cells exocytosis now occurs at the front of the cell: It is here that both LDL- and transferrin-receptors emerge from inside the cell and return to the cell surface. As the sites of endocytosis (at random on the cell surface) and exocytosis (at the front edge of the cell) are separated in space, it follows that (within the cell’s context) there is a flow of membrane from the front rearward.
The consequences of this polarised endocytic cycle are profound:
• The membrane added at the front of the cell is believed to provide the surface there for the cell to extend itself forward as it moves.
• Evidence indicates that those molecules in the cell’s surface that act as the feet of the cell — the integrins, which attach the cell to the substratum — can also be endocytosed and transported through the cell. In this way, fresh adhesion sites (see cell adhesion) are provided at the cell’s front.
• The flow of membrane from the front rearward is not a flow of average plasma membrane proteins: The membrane internalised is a subset of plasma membrane proteins, such as LDL or transferrin receptors and the lipid bilayer in which they sit. These recycling molecules appear at the front surface of the cell and diffuse about, drifting rearwards until they are recaptured by a coated pit and transported back, through the cell, to the front. Other proteins do not participate in this cycle: Therefore, they experience a slow rearward flow of the lipid bilayer in which they reside. Thus, they are subject to two different influences: (a) they tend to get swept backward in this lipid flow and (b) they tend to randomise their distribution on the cell surface by Brownian motion. Calculation shows that, for non-cycling proteins, diffusion is the more important influence, so these molecules would be expected to have a near-random distribution on the cell’s surface. However, if a large object were attached to the cell surface that were unable to diffuse against the flow, it would be expected to be swept backward by the flow toward the trailing end of the cell. Indeed, it would act as a marker for that flow. This is the reason why carbon particles attached to the dorsal surface of a moving cell, or why aggregates of surface proteins, are seen to move to the back of a cell. This process is known as cap formation.
Further support for this scheme comes from a study of yeast cells (S. cerevisiae), which undergo a primitive form of movement called shmooing (after Al Capp's shmoo). A protein that cycles rapidly would be expected to be most concentrated near the shmoo tip of such a cell, whereas a non-cycling protein would be expected to be swept backward slightly. Both these distributions have been observed for a cycling and non-cycling variant of the same protein in the plasma membrane of yeast cells as they shmoo.
• The molecular feet of a cell (usually integrins), when bound to the substratum, cannot diffuse about. Like any other macroscopic object sitting in the lipid flow, they are thus pushed backward. However, they cannot move backward (they are attached to the fixed substratum)) and therefore push the cell forward. This may be the force against the substratum, which enables a cell to move forward.
• Polarised endocytic cycles are believed to exist in other cellular contexts, but the evidence for them at present is less clear.