
 | ||
Cryo-electron microscopy (cryo-EM), or electron cryomicroscopy, is a form of transmission electron microscopy (TEM) where the sample is studied at cryogenic temperatures (generally liquid-nitrogen temperatures). Cryo-EM is gaining popularity in structural biology.
Contents
- Development
- Thin film
- Vitreous sections
- Material specimens
- Techniques in cryoelectron microscopy
- References
The utility of cryoelectron microscopy stems from the fact that it allows the observation of specimens that have not been stained or fixed in any way, showing them in their native environment. This is in contrast to X-ray crystallography, which requires crystallizing the specimen, which can be difficult, and placing them in non-physiological environments, which can occasionally lead to functionally irrelevant conformational changes.
The resolution of cryo-EM maps is improving steadily, and in 2014 some structures at near-atomic resolution had been obtained using cryoelectron microscopy, including those of viruses, ribosomes, mitochondria, ion channels, and enzyme complexes as small as 170 kD at a resolution of 4.5 Å. Bridget Carragher and her colleagues at the Scripps National Resource for Automated Molecular Microscopy used techniques she and Clint Potter developed to create the first cryo-electron microscopy structural biology image with a resolution finer than 3 angstroms, thereby elevating cryo-EM as a tool comparable to and potentially superior to traditional x-ray crystallography techniques. A 2.2 Å map of a bacterial enzyme beta-galactosidase was published in June 2015. A version of electron cryomicroscopy is cryo-electron tomography (CET), where a 3D reconstruction of a sample is created from tilted 2D images.
Development
The original rationale for cryoelectron microscopy was as a means to fight radiation damage for biological specimens. The amount of radiation required to collect an image of a specimen in the electron microscope is high enough to be a potential source of specimen damage for delicate structures. In addition, the high vacuum required on the column of an electron microscope makes the environment for the sample quite harsh.
The problem of the vacuum was partially solved by the introduction of negative stains but even with negative stains biological samples are prone to structural collapse upon dehydration of the specimen. Embedding the samples in ice below the sublimation temperature was a possibility that was contemplated early on, but water tends to arrange into a crystalline lattice of lower density upon freezing and this can destroy the structure of anything that is embedded in it.
In the early '80s, several groups studying solid state physics were attempting to produce vitreous ice by different means, such as high pressure freezing or flash freezing. In a seminal paper in 1984, the group led by Jacques Dubochet at the European Molecular Biology Laboratory showed images of adenovirus embedded in a vitrified layer of water. This paper is generally considered to mark the origin of cryoelectron microscopy, and the technique has been developed to the point of becoming routine at numerous laboratories throughout the world.
The energy of the electrons used for imaging (80-300 kV) is high enough that covalent bonds can be broken. When imaging specimens vulnerable to radiation damage, it is necessary to limit the electron exposure used to acquire the image. These low exposures require that the images of thousands or even millions of identical frozen molecules be selected, aligned, and averaged to obtain high-resolution maps, using specialized software. A significant improvement in structural features was achieved in 2012 by the introduction of direct electron detectors and better computational algorithms.
Thin film
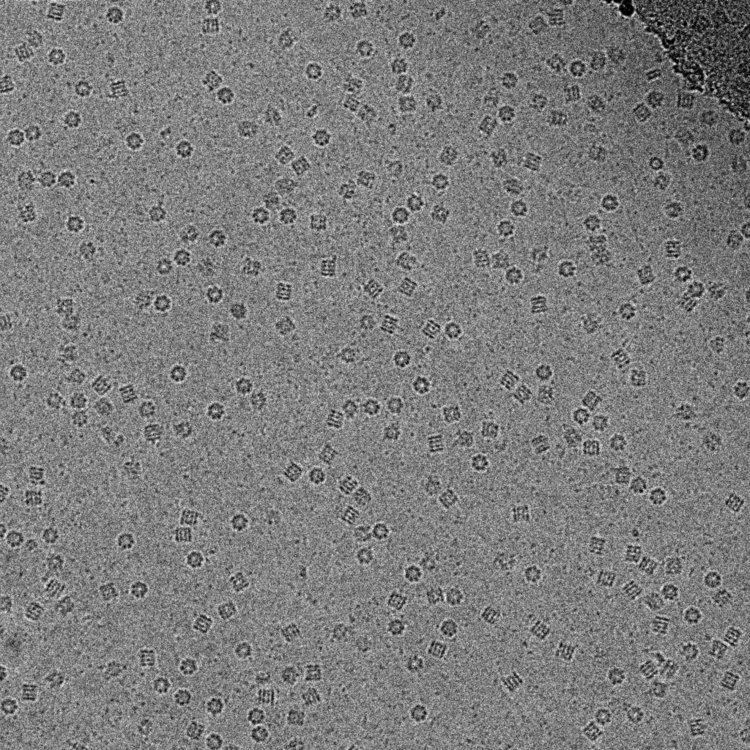
The biological material is spread on an electron microscopy grid and is preserved in a frozen-hydrated state by rapid freezing, usually in liquid ethane near liquid nitrogen temperature. By maintaining specimens at liquid nitrogen temperature or colder, they can be introduced into the high-vacuum of the electron microscope column. Most biological specimens are extremely radiation sensitive, so they must be imaged with low-dose techniques (usefully, the low temperature of cryo-electron microscopy provides an additional protective factor against radiation damage).
Consequently, the images are extremely noisy. For some biological systems it is possible to average images to increase the signal-to-noise ratio and retrieve high-resolution information about the specimen using the technique known as single particle analysis. This approach in general requires that the things being averaged are identical, although some limited conformational heterogeneity can now be studied (e.g. ribosome). Three-dimensional reconstructions from cryo-EM images of protein complexes and viruses have been solved to sub-nanometer or near-atomic resolution, allowing new insights into the structure and biology of these large assemblies.
Analysis of ordered arrays of protein, such as 2-D crystals of transmembrane proteins or helical arrays of proteins, also allows a kind of averaging which can provide high-resolution information about the specimen. This technique is called electron crystallography.
Vitreous sections
The thin film method is limited to thin specimens (typically < 500 nm) because the electrons cannot cross thicker samples without multiple scattering events. Thicker specimens can be vitrified by plunge freezing (cryofixation) in ethane (up to tens of μm in thickness) or more commonly by high pressure freezing (up to hundreds of μm). They can then be cut in thin sections (40 to 200 nm thick) with a diamond knife in a cryoultramicrotome at temperatures lower than -135 °C (devitrification temperature). The sections are collected on an electron microscope grid and are imaged in the same manner as specimen vitrified in thin film. This technique is called cryo-electron microscopy of vitreous sections (CEMOVIS) or cryo-electron microscopy of frozen-hydrated sections.
Material specimens
In addition to allowing vitrified biological samples to be imaged, cryo-EM can also be used to image material specimens that are too volatile in vacuum to image using standard, room temperature electron microscopy. For example, vitrified sections of liquid-solid interfaces can be extracted for analysis by cryo-EM, and sulfur, which is prone to sublimation in the vacuum of electron microscopes, can be stabilized and imaged in cryo-EM.
Techniques in cryoelectron microscopy
A variety of techniques can be used in cryoelectron microscopy. Popular techniques include:
- Electron crystallography
- Analysis of two-dimensional crystals
- Analysis of helical filaments or tubes
- Single particle analysis
- Cryo-electron tomography
- MicroED