
 | ||
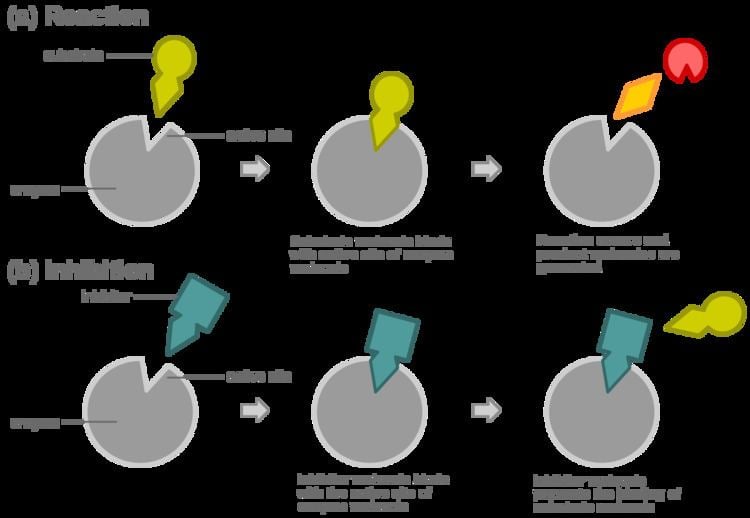
Competitive inhibition is a form of enzyme inhibition where binding of the inhibitor to the active site on the enzyme prevents binding of the substrate and vice versa.
Contents
Most competitive inhibitors function by binding reversibly to the active site of the enzyme. As a result, many sources state that this is the defining feature of competitive inhibitors. This, however, is a misleading oversimplification, as there are many possible mechanisms by which an enzyme may bind either the inhibitor or the substrate but never both at the same time. For example, allosteric inhibitors may display competitive, non-competitive, or uncompetitive inhibition.
Mechanism
In competitive inhibition, at any given moment, the enzyme may be bound to the inhibitor, the substrate, or neither, but it cannot bind both at the same time.
In virtually every case, competitive inhibitors bind in the same binding site as the substrate, but same-site binding is not a requirement. A competitive inhibitor could bind to an allosteric site of the free enzyme and prevent substrate binding, as long as it does not bind to the allosteric site when the substrate is bound. For example, strychnine acts as an allosteric inhibitor of the glycine receptor in the mammalian spinal cord and brain stem. Glycine is a major post-synaptic inhibitory neurotransmitter with a specific receptor site. Strychnine binds to an alternate site that reduces the affinity of the glycine receptor for glycine, resulting in convulsions due to lessened inhibition by the glycine.
In competitive inhibition, the maximum velocity (
Equation
Competitive inhibition increases the apparent value of the Michaelis-Menten constant,
where
Derivation
In the simplest case of a single-substrate enzyme obeying Michaelis-Menten kinetics, the typical scheme
is modified to include binding of the inhibitor to the free enzyme:
Note that the inhibitor does not bind to the ES complex and the substrate does not bind to the EI complex. It is generally assumed that this behavior is indicative of both compounds binding at the same site, but that is not strictly necessary. As with the derivation of the Michaelis-Menten equation, assume that the system is at steady-state, i.e. the concentration of each of the enzyme species is not changing.
Furthermore, the known total enzyme concentration is
We can therefore set up a system of equations:
where
From equation (3), we can define E in terms of ES by rearranging to
Dividing by
As in the derivation of the Michaelis-Menten equation, the term
Substituting equation (5) into equation (4), we have
Rearranging, we find that
At this point, we can define the dissociation constant for the inhibitor as
At this point, substitute equation (5) and equation (6) into equation (1):
Rearranging to solve for ES, we find
Returning to our expression for
Since the velocity is maximal when all the enzyme is bound as the enzyme-substrate complex,
To compute the concentration of competitive inhibitor