
 | ||
Neural crest cells are a group of temporary, multipotent (can give rise to some other types of cells but not all) cells that are pinched off during the formation of the neural tube (precursor to the spinal cord and brain) and therefore are found at the dorsal (top) region of the neural tube during development. They are derived from the ectoderm germ layer, but are sometimes called the fourth germ layer because they are so important and give rise to so many other types of cells. They migrate throughout the body and create a large number of differentiated cells such as neurons, glial cells, pigment-containing cells in skin, skeletal tissue cells in the head, and many more.
Contents
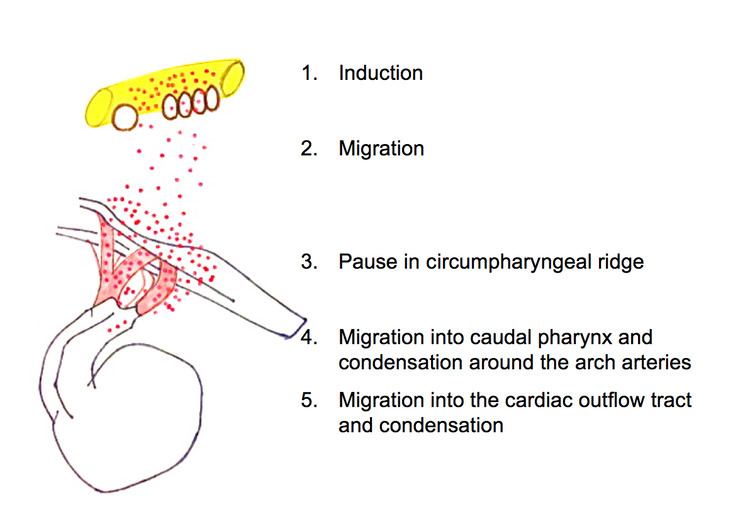
- Induction
- Initial migration
- Pause at the circumpharyngeal ridge
- Migration to the pharyngeal arches
- Migration to the cardiac outflow tract
- Molecular pathways
- Wnt
- Notch
- Bone morphogenetic proteins
- Fibroblast growth factor 8
- GATA
- CNCCS and ischaemic heart disease
- References
Cardiac neural crest cells (CNCCs) are a type of neural crest cells that migrate to the circumpharyngeal ridge (an arc-shape ridge above the pharyngeal arches) and then into the 3rd, 4th and 6th pharyngeal arches and the cardiac outflow tract (OFT).
They extend from the otic placodes (the structure in developing embryos that will later form the ears) to the third somites (clusters of mesoderm that will become skeletal muscle, vertebrae and dermis).
The cardiac neural crest cells have a number of functions including creation of the muscle and connective tissue walls of large arteries; parts of the cardiac septum; parts of the thyroid, parathyroid and thymus glands. They differentiate into melanocytes and neurons and the cartilage and connective tissue of the pharyngeal arches. They may also contribute to the creation of the carotid body, the organ which monitors oxygen in the blood and regulates breathing.
Induction
Induction is the differentiation of progenitor cells into their final designation or type. The progenitor cells which will become CNCCs are found in the epiblast about Henson’s node. Progenitor cells are brought into the neural folds. Molecules such as Wnt, fibroblast growth factor (FGF) and bone morphogenetic protein (BMP) provide signals which induce the progenitor cells to become CNCCs. Little is known about the signal cascade that promotes neural crest induction. However, it is known that an intermediate level of BMP is required: if BMP is too high or too low, the cells do not migrate.
Initial migration
After induction, CNCCs lose their cell to cell contacts. This allows them to move through the extracellular matrix and interact with its components. The CNCCs, with the assistance of their filopodia and lamellipodia (actin containing extensions of cytoplasm that allow a cell to probe its path of migration), leave the neural tube and migrate along a dorsolateral pathway to the circumpharyngeal ridge. Along this pathway, CNCCs link together to form a stream of migrating cells. Cells at the front of the migration stream have a special polygonal shape and proliferate at a faster rate than trailing cells.
Pause at the circumpharyngeal ridge
At the circumpharyngeal arch the CNCCs must pause in their migration while the pharyngeal arches form.
Migration to the pharyngeal arches
The CNCCs continue their migration into the newly formed pharyngeal arches, particularly the third, fourth and sixth arches. In the pharyngeal arches the CNCCs assist in the formation of the thyroid and parathyroid glands.
The leading cells have long filopodia that assist migration while cells in the middle of the migration have protrusions at their front and back allowing them to interact and communicate with leading cells, trailing cells and receive signals from the extracellular matrix.
A variety of growth factors and transcription factors in the extracellular matrix signal cells and direct them toward a specific arch. For example, signalling by FGF8 directs CNCCS to the fourth arch and keeps the cells viable.
Migration to the cardiac outflow tract
The cardiac outflow tract is a temporary structure in the developing embryo that connects the ventricles with the aortic sac. Some CNCCs migrate beyond the pharyngeal arches to the cardiac outflow tract. CNCCS in the cardiac outflow tract contribute to the formation of the cardiac ganglia and mesenchyme at the junction of the subaortic and sub pulmonary myocardium (muscular heart tissue) of the outflow tract. A smaller portion of the CNCCs migrate to the proximal outflow tract where they help to close the ventricular outflow septum.
Molecular pathways
Many signaling molecules are required for the differentiation, proliferation, migration and apoptosis of the CNCCs. The molecular pathways involved include the Wnt, Notch, BMP, FGF8 and GATA families of molecules. In addition to these signaling pathways, these processes are also mediated by environmental factors including blood flow, shear stress, and blood pressure.
The CNCCs interact with the cardiogenic mesoderm cells of the primary and secondary heart fields, which are derived from the cardiac crescent and will give rise to the endocardium, myocardium, and epicardium. The CNCCs themselves are the precursors to vascular smooth muscle cells and cardiac neurons.
For example, CNCCs are required for the formation of the aorticopulmonary septum (APS) that directs cardiac outflow into two tracts: the pulmonary trunk and the aorta of the developing heart. This is an example of remodelling which is dependent on signalling back and forth between CNCCs and the cardiogenic mesoderm. If this signalling is disrupted or there are defects in the CNCCS, cardiovascular anomalies may develop. These anomalies include persistent truncus arteriosus (PTA), double outlet right ventricle (DORV), tetralogy of Fallot and DiGeorge syndrome.
Wnt
Wnt proteins are extracellular growth factors that activate intracellular signalling pathways. There are two types of pathways: canonical and non-canonical. The classic canonical Wnt pathway involves B-catenin protein as a signaling mediator. Wnt maintains B-catenin by preventing against Proteasome degradation. Thus, B-catenin is stabilized in the presence of Wnt and regulates gene transcription through interaction with TCF/LEF transcription factors. The canonical Wnt/B-catenin pathway is important for control of cell proliferation. The non-canonical Wnt pathway is independent of B-catenin and has an inhibitory effect on canonical Wnt signaling.
Wnt signaling pathways play a role in CNCC development as well as OFT development. In mice, decrease of B-catenin results in a decrease in the proliferation of CNCCs. Downregulation of the Wnt coreceptor Lrp6 leads to a reduction of CNCCs in the dorsal neural tube and in the pharyngeal arches, and results in ventricular, septal, and OFT defects. Canonical Wnt signaling is especially important for cell cycle regulation of CNCC development and the initiation of CNCC migration. Non-canonical Wnt signaling plays a greater role in promoting cardiac differentiation and OFT development.
Notch
Notch is a transmembrane protein whose signaling is required for differentiation of CNCCs to vascular smooth muscle cells and for proliferation of cardiac myocytes (muscle cells of the heart). In mice, disruption of Notch signaling results in aortic arch branching defects and pulmonary stenosis, as well as a defect in the development of the smooth muscle cells of the sixth aortic arch artery, which is the precursor to the pulmonary artery. In humans, mutations in Notch most often result in bicuspid aortic valve disease and calcification of the aortic valve.
Bone morphogenetic proteins
Bone morphogenetic proteins (BMPs) are required for neural crest cell migration into the cardiac cushions (precursors to heart valves and septa) and for differentiation of neural crest cells to smooth muscle cells of the aortic arch arteries. In neural crest–specific Alk2-deficient embryos, the cardiac cushions of the outflow tract are deficient in cells because of defects in neural crest cell migration.
Fibroblast growth factor 8
Fibroblast growth factor 8 (FGF8) transcription factors are essential for regulating the addition of secondary heart field cells into the cardiac outflow tract. FGF8 mouse mutants have a range of cardiac defects including underdeveloped arch arteries and transposition of the great arteries.
GATA
GATA transcription factors, which are complex molecules that bind to the DNA sequence GATA, play a critical role in cell lineage differentiation restriction during cardiac development. The primary function of GATA6 in cardiovascular development is to regulate the morphogenetic patterning of the outflow tract and aortic arch. When GATA6 is inactivated in CNCCs, various cardiovascular defects such as persistent truncus arteriorus and interrupted aortic arch may occur. This phenotype (anomaly) was also observed when GATA6 was inactivated within vascular smooth muscle cells. GATA6 in combination with Wnt (Wnt2-GATA6) plays a role in the development of the posterior pole of the heart (the inflow tract).
CNCCS and ischaemic heart disease
There is interest amongst researchers as to whether CNCCs can be used to repair human heart tissue. Heart attacks in humans are common and their rate of mortality is high. There are emergency treatments that hospitals can administer, such as angioplasty or surgery, but after that patients will likely be on medication for the long term and are more susceptible to heart attacks in the future. Other complications of heart attacks include cardiac arrhythmias and heart failure.
Although CNCCs are important in embryos, some CNCCs are retained in a dormant state to adulthood where they are called neural crest stem cells. In 2005, Tomita transplanted neural crest stem cells from mammal hearts to the neural crest of chick embryos. These CNCCs were shown to migrate into the developing heart of the chick using the same dorsolateral pathway as the CNCCs, and differentiate into neural and glial cells.
Tamura's study of 2011 examined the fate of CNCCs after a heart attack (myocardial infarction) in young mice. The CNCCs in the young mice were tagged with enhanced green fluorescent protein (EGFP) and then traced. Tagged CNCCs were concentrated in the cardiac outflow tract, and some were found in the ventricular myocardium. These cells were also shown to be differentiating into cardiomyocytes as the heart grew. Although less were found, these EGFP-labelled CNCCs were still present in the adult heart. When a heart attack was induced, the CNCCs aggregated in the ischemic border zone area (an area of damaged tissue that can still be saved) and helped contribute to the regeneration of the tissue to some extent via differentiation into cardiomyocytes to replace the necrotic tissue.