
Domain Eukaryota Scientific name Blepharisma Higher classification Blepharismidae Order Heterotrich | Family Blepharismidae Rank Genus | |
 | ||
Similar Blepharisma japonicum, Stentor, Heterotrich, Spirostomum, Hypotrich | ||
Blepharisma mpg
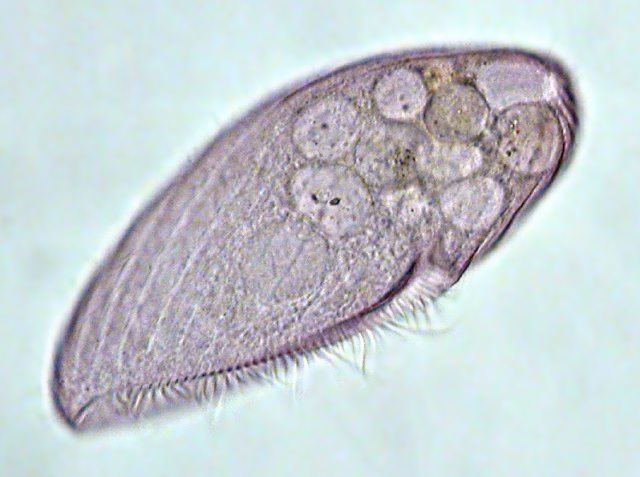
Blepharisma is a genus of unicellular ciliate protists found in fresh and salt water. The group includes about 40 accepted species, and many sub-varieties and strains. While species vary considerably in size and shape, most are easily identified by their red or pinkish color, which is caused by granules of the pigment Blepharismin.
Contents
- Blepharisma mpg
- Blepharisma friends
- Appearance
- Reproduction
- Feeding and behavior
- List of Species
- References
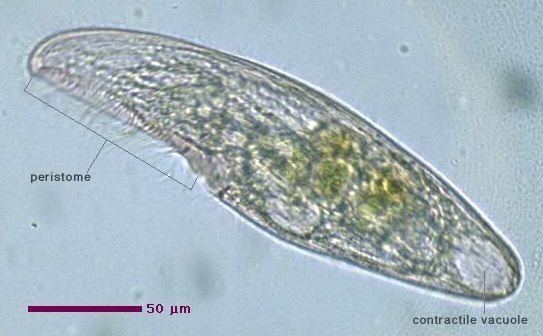
All members of the genus possess a long series of membranelles on the left side of the oral groove, and an "undulating membrane" (a structure resembling a flap, scarf or small sail, composed of long cilia fused together as a single sheet) on the right side of the peristome, toward the posterior.
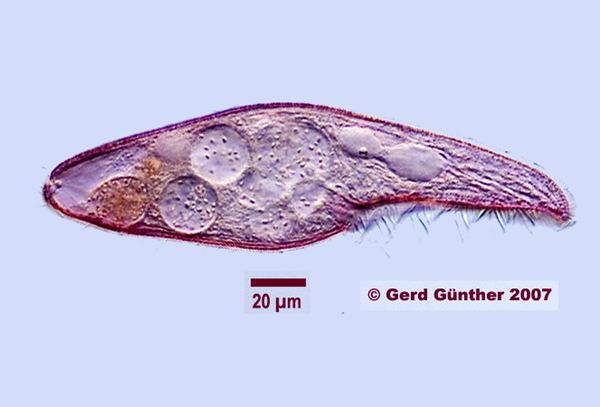
Certain species of Blepharisma have served as model organisms for scientific research. Since some varieties are easily cultured and readily available from scientific supply houses, they are a common object of study in school science classes.
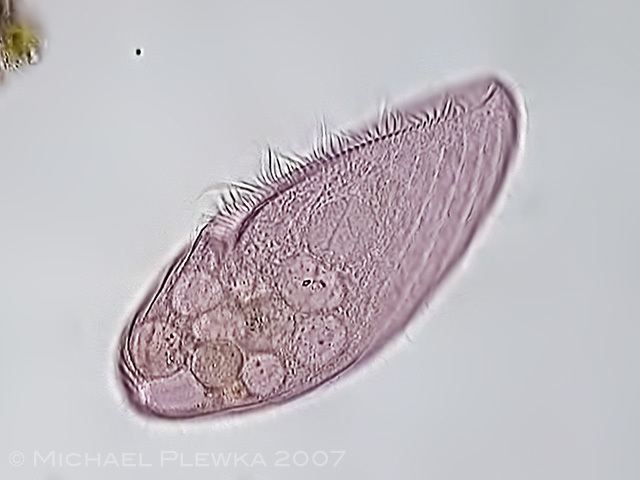
Blepharisma friends
Appearance
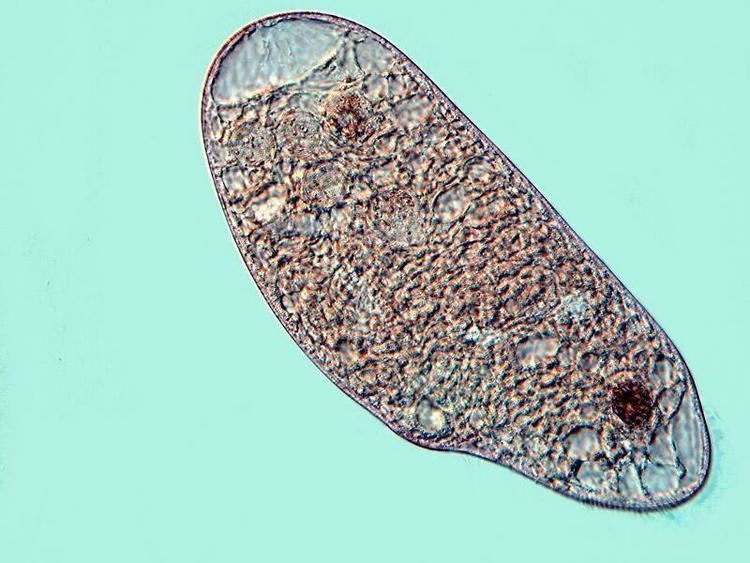
Blepharisma may be as small as 50 micrometres in length, or as large as 1 mm (though normal size range is between 75 and 300 micrometres). Body shape varies within the genus. The type species of the genus, B. persicinum, is ellipsoidal. Blepharisma lateritium is teardrop-shaped, with a rounded posterior; while Blepharisma elongatum and Blepharisma sphagni are long and thin, tapering at the posterior into a tail-like point.
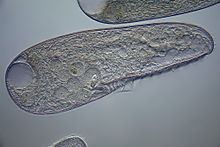
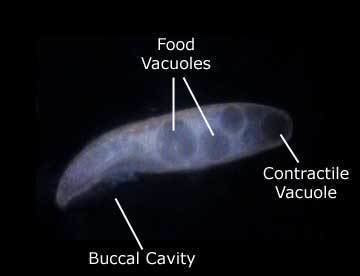
All species are uniformly ciliated, with the cilia arranged in longitudinal rows, and stripes of pigment alternating between rows of cilia. The pink or red pigmentation may be quite pale, and in certain cases it is absent altogether. A contractile vacuole, often quite large, is located in the posterior. The Macronuclei can take a variety of forms. Depending on species and phase of life, they may be rod-shaped, ovoid, spherical, or moniliform (like a rosary, or string of beads).
Reproduction
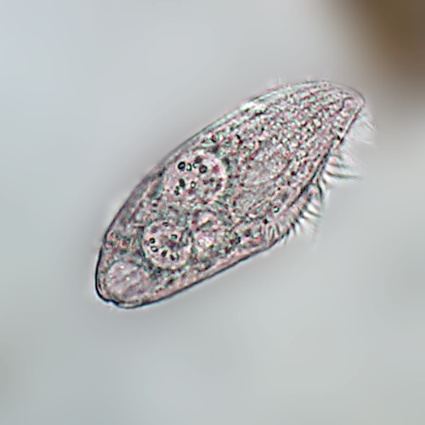
Like all ciliates, Blepharisma can reproduce by binary fission, dividing transversally. They also engage in a type of sexual reproduction known as ciliate conjugation, a process through which genetic material can pass directly from one cell to the other. In conjugation, two organisms come into close contact, and a temporary cytoplasmic bridge forms between them. The micronuclei of each cell then undergo meiosis, and haploid micronuclei pass from one individual to the other. This permits the reshuffling of hereditary characteristics, as in other types of sexual reproduction. Conjugation is immediately followed by binary fission of the two conjugants.
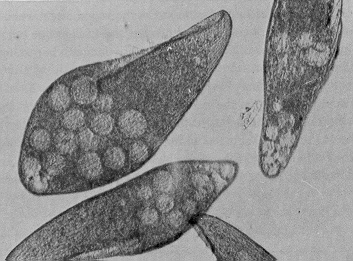
In Blepharisma, as in some other ciliates, chemical substances called gamones are used to induce conjugation by stimulating interaction between compatible mating partners.
Although Blepharisma are able to undergo a form of sexual reproduction that involves selfing, ordinarily sexual reproduction involves interaction of cells of different mating types. There are two mating types (I and II), each type excreting a specific pheromone (termed gamone 1 and gamone 2, respectively). When sexually mature mating-type I cells are moderately starved, they autonomously produce and secrete gamone I. Gamone 1 specifically acts on mating-type II cells, transforming them so that they can unite with type I cells, and inducing them to secrete gamone 2. Gamone 2 then transforms type I cells so that they can unite with type II cells. Cells that can unite may then undergo conjugation. Sexual reproduction involving interaction of opposite mating types promotes outcrossing and the masking of deleterious recessive mutations in the diploid phase of the sexual cycle.
Feeding and behavior
Blepharisma feed on a variety of smaller organisms, including bacteria, flagellate algae, rotifers, other ciliates and even smaller members of the same species. Experiments with Blepharisma undulans have shown that cannibalism causes gigantism. When individuals are given a diet of smaller Blepharisma, or certain ciliates (particularly Colpidium colpoda), they grow to a relatively enormous size. As long as their diet remains unchanged, cannibal giants will divide to produce more giants. When large prey become unavailable, the offspring will revert to normal size.
Blepharisma are markedly photophobic, and when light levels are increased will seek out darkened areas. The ability to detect light is accomplished with photosensitive pigment granules located just under the plasma membrane of the cell. The pigment in these granules is Blepharismin, the same substance that gives Blepharisma their characteristic pinkish color.