
 | ||
Bacterial circadian rhythms, like other circadian rhythms, are endogenous "biological clocks" that have the following three characteristics: (a) in constant conditions (i.e. constant temperature and either constant light {LL} or constant darkness {DD}) they oscillate with a period that is close to, but not exactly, 24 hours in duration, (b) this "free-running" rhythm is temperature compensated, and (c) the rhythm will entrain to an appropriate environmental cycle.
Contents
- History are prokaryotes capable of circadian rhythmicity
- Relationship to cell division
- Adaptive significance
- Global regulation of gene expression and chromosomal topology
- Molecular mechanism of the cyanobacterial clockwork
- Visualizing the clockworks gears structural biology of clock proteins
- References
Until the mid-1980s, it was thought that only eukaryotic cells had circadian rhythms. It is now known that cyanobacteria (a phylum of photosynthetic eubacteria) have well-documented circadian rhythms that meet all the criteria of bona fide circadian rhythms. In these bacteria, three key proteins whose structures have been determined can form a molecular clockwork that orchestrates global gene expression. This system enhances the fitness of cyanobacteria in rhythmic environments.
History: are prokaryotes capable of circadian rhythmicity?
Before the mid-1980s, it was believed that only eukaryotes had circadian systems. The conclusion that only eukaryotes have circadian oscillators seemed reasonable, because it was assumed that an endogenous timekeeper with a period close to 24 hours would not be useful to prokaryotic organisms that often divide more rapidly than once every 24 hours. The assumption might be stated as, "why have a timer for a cycle that is longer than your lifetime?" While intuitive, the conclusion was flawed. It was based on the assumption that a bacterial cell is equivalent to a sexually reproducing multicellular organism. However, a bacterial culture is more like a mass of protoplasm that grows larger and larger and incidentally subdivides. From this perspective, it is reasonable that a 24-hour temporal program could be adaptive to a rapidly dividing protoplasm if the fitness of that protoplasm changes as a function of daily alterations in the environment (light intensity, temperature, etc.).
In 1985–86, several research groups discovered that cyanobacteria display daily rhythms of nitrogen fixation in both light/dark (LD) cycles and in constant light. The group of Huang and co-workers was the first to recognize clearly that the cyanobacterium Synechococcus sp. RF-1 was exhibiting circadian rhythms, and in a series of publications beginning in 1986 demonstrated all three of the salient characteristics of circadian rhythms described above in the same organism, the unicellular freshwater Synechococcus sp. RF-1. Another ground-breaking study was that of Sweeney and Borgese, who were the first to demonstrate temperature compensation of a daily rhythm in the marine cyanobacterium, Synechococcus WH7803.
Inspired by the research of the aforementioned pioneers, the cyanobacterium Synechococcus elongatus was genetically transformed with a luciferase reporter that allowed rhythmic gene expression to be assayed non-invasively as rhythmically "glowing" cells. This system allowed an exquisitely precise circadian rhythm of luminescence to be measured from cell populations and even from single cyanobacterial cells. The luminescence rhythms expressed by these transformed S. elongatus fulfilled all three key criteria of circadian rhythms: persistence of a 24-hour oscillation in constant conditions, temperature compensation, and entrainment. Thus, the work with various Synechococcus species firmly established that prokaryotic bacteria are capable of circadian rhythmicity, displacing the prior "no circadian clocks in prokaryotes" dogma. Nevertheless, persuasive evidence for circadian programs in bacteria other than the cyanobacteria is still lacking.
Relationship to cell division
Despite predictions that circadian clocks would not be expressed by cells that are doubling faster than once per 24 hours, the cyanobacterial rhythms continue in cultures that are growing with doubling times as rapid as one division every 5–6 hours. Apparently cyanobacteria are able to simultaneously and accurately keep track of two timing processes that express significantly different periods.
Adaptive significance
Do circadian timekeepers enhance the fitness of organisms growing under natural conditions? Despite the expectation that circadian clocks are usually assumed to enhance the fitness of organisms by improving their ability to adapt to daily cycles in environmental factors, there have been few rigorous tests of that proposition in any organism. Cyanobacteria are one of the few organisms in which such a test has been performed. The adaptive fitness test was done by mixing cyanobacterial strains that express different circadian properties (i.e., rhythmicity vs. arhythmicity, different periods, etc.) and growing them in competition under different environmental conditions. The idea was to determine if having an appropriately functional clock system enhances fitness under competitive conditions. The result was that strains with a functioning biological clock out-compete arhythmic strains in environments that have a rhythmic light/dark cycle (e.g., 12 hours of light alternating with 12 hours of darkness), whereas in "constant" environments (e.g., constant illumination) rhythmic and arhythmic strains grow at comparable rates. Among rhythmic strains with different periods, the strains whose endogenous period most closely matches the period of the environmental cycle is able to out-compete strains whose period does not match that of the environment. Therefore, in rhythmic environments, the fitness of cyanobacteria is improved when the clock is operational and when its circadian period is similar to the period of the environmental cycle. These were among the first rigorous demonstrations in any organism of a fitness advantage conferred by a circadian system.
When researchers believed that prokaryotes were too "simple" to have circadian timekeepers, it had seemed reasonable that evolutionary selection for circadian organization would occur only if the generation time of the cells were as long or longer than a day. The data from cyanobacteria, however, suggest that the benefits of having a daily clock can potentially accrue to all organisms, even if they divide more rapidly than once a day.
Global regulation of gene expression and chromosomal topology
In eukaryotes, about 10–20% of the genes are rhythmically expressed (as gauged by rhythms of mRNA abundance). However, in cyanobacteria, a much larger percentage of genes are controlled by the circadian clock. For example, one study has shown that the activity of essentially all promoters are rhythmically regulated. The mechanism by which this global gene regulation is mechanistically linked to the circadian clock is not known, but it may be related to rhythmic changes in the topology of the entire cyanobacterial chromosome.
Molecular mechanism of the cyanobacterial clockwork
The S. elongatus luciferase reporter system was used to screen for clock gene mutants, of which many were isolated. These mutants were used to identify a gene cluster composed of three genes, named kaiA, kaiB and kaiC; (Ishiura et al., 1998; "kai" means "rotation" or "cycle number" in Japanese). These genes encode the proteins KaiA, KaiB, and KaiC, which are essential for clock function in S. elongatus and constitute a core circadian oscillator. No significant similarity was found among the kai genes and any other previously reported genes in eukaryotes, but there are potential homologs in the genomic sequences of other bacteria (both eubacteria and archaea).
At first, the cyanobacterial clockwork appeared to be a transcription and translation feedback loop in which clock proteins autoregulate the activity of their own promoters by a process that was similar in concept to the circadian clock loops of eukaryotes. Subsequently, however, several lines of evidence indicated that transcription and translation was not necessary for circadian rhythms of Kai proteins, the most spectacular being that the three purified Kai proteins can reconstitute a temperature-compensated circadian oscillation in a test tube. The rhythm that is measurable in vitro is the phosphorylation status of the clock protein KaiC. This is the first (and so far, only) example of the reconstitution of a circadian clock in vitro.
Visualizing the clockwork's "gears": structural biology of clock proteins
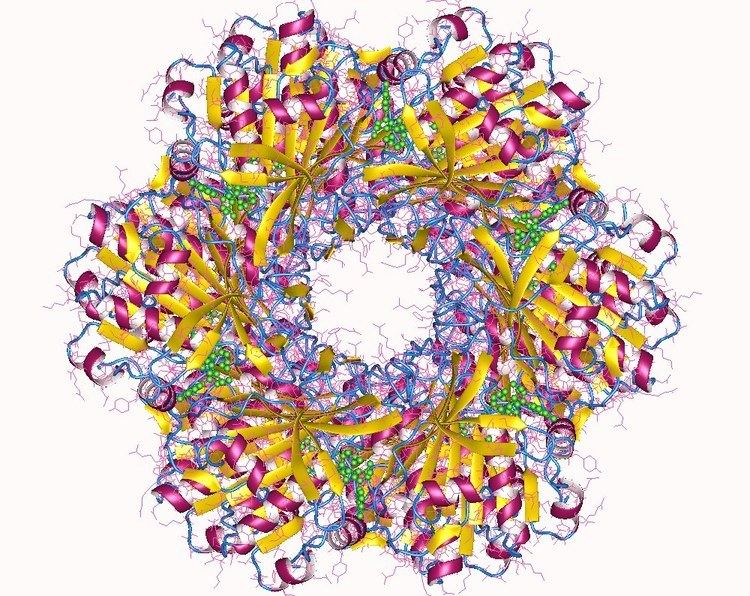
The cyanobacterial circadian system is so far unique in that it is the only circadian system in which the structures of full-length clock proteins have been solved. In fact, the structures of all three of the Kai proteins have been determined. KaiC forms a hexamer that resembles a double doughnut with a central pore that is partially sealed at one end. There are twelve ATP-binding sites in KaiC and the residues that are phosphorylated during the in vitro phosphorylation rhythm have been identified. KaiA has two major domains and forms dimers in which the N-terminal domains are "swapped" with the C-terminal domains. KaiB has been successfully crystallized from three different species of cyanobacteria and forms dimers or tetramers.
The three-dimensional structures have been helpful in elucidating the cyanobacterial clock mechanism by providing concrete models for the ways in which the three Kai proteins interact and influence each other. The structural approaches have also allowed the KaiA/KaiB/KaiC complexes to be visualized as a function of time, which enabled sophisticated mathematical modeling of the in vitro phosphorylation rhythm. Therefore, the cyanobacterial clock components and their interactions can be visualized in four dimensions (three in space, one in time).